
Замкнутые нейронные цепи. Интересно, что основой для гипотезы о физиологическом механизме кратковременной памяти послужили морфологические данные. Форбс (Forbes) в 1920 г. сообщил, что в центральной нервной системе наряду с разомкнутыми нейронными цепями на всех уровнях существуют также сложные замкнутые сети. Эти наблюдения использовал Лоренте де Но, который дал точное описание таких сетей в разных частях больших полушарий. На основе этого морфоло- ческого описания физиологам было уже нетрудно предположить практически нескончаемую реверберацию импульсов, не требующую подкрепления новыми сенсорными стимулами. Эти самостимулируемые, так называемые реверберирующие, замкнутые цепи, возможно, лежат в основе кратковременной памяти (рис. 53).
Самостимулируемая цепь. Базируясь на морфологических данных, Рашевски (Rashevsky) создал в 1938 г. модель памяти, состоящую из замкнутых цепей, в которых импульсы долгое время циркулируют без подкрепления. Новые афферентные импульсы только усиливают поток циркулирующих импульсов. Рашевски предположил также, что эти цепи играют роль не только в кратковременной памяти, но и в выработке условных рефлексов. Импульсация, вызываемая новым, условным стимулом, суммируется с реверберирующей волной деполяризации, вызывая тем самым условную реакцию. Рашевски дал также математическое описание своей теории, которая и поныне служит отправным пунктом современных теорий реверберационно- го хранения информации. Он применил свою математическую модель к различным психологическим процессам, связанным с памятью, но не смог объяснить с помощью этой теории усиление долговременной памяти со временем, особенно в старости.
Существование самостимулируемых цепей в сером веществе головного мозга было доказано только в 60-х годах Верцеано и Негиси, которые вводили микроэлектроды в различные нейроны, лежащие на небольшом пространстве в несколько квадратных миллиметров. Они наблюдали волну импульсов, вызванную стимуляцией, которая с некоторой задержкой переходила от клетки к клетке. Задержка соответствовала времени, необходимому для синаптической передачи.
Электрическая модель памяти. В наших экспериментах, начатых также в 60-х годах, в коре, таламусе и ретикулярной формации среднего мозга у кошки регистрировались условные вызванные потенциалы при электрическом раздражении аффе-
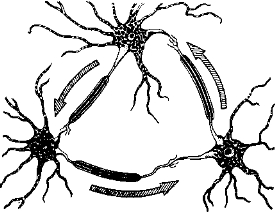
Рис. 53. Схема реверберирующей нейронной цепи.
рентных нервов. Эти выработанные вызванные потенциалы мы рассматривали как «электрическую модель памяти». Без подкрепления ответы, возникавшие в одном опыте, быстро исчезали. Таким образом, их можно было считать следами кратковременной, лабильной памяти, основанными на реверберирующей волне потенциалов действия в самостимулируемой цепи соответствующей группы мозговых нейронов. Для подтверждения нашей гипотезы был применен электрошок.
Действие электрошока. Воздействие электрошоком, введенное Черлетти и Бини (Cerletti, Bini), вызвало в литературе много споров. Действие на мозг электрического импульса в 120 В и 120—500 мА в течение 0,5—1 с вызывает эпилептоид- ные судороги, Такой электрошок приводит к полной потере сознания приблизительно на 3 мин. В течение 30 мин после шока наблюдается постепенно исчезающее затемнение памяти. Судороги длятся около минуты (тоническая фаза — несколько секунд, а затем клоническая фаза в течение 30 с). Согласно большинству авторов, импульс высокочастотного тока вызывает
Контроль
До шона
После
шока
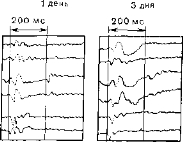
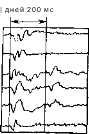
Рнс. 54. Вверху: развитие запаздывающего вызванного потенциала у кошки. Внизу: спустя один день выработанный ответ еще можно угасить электрошоком, но через три и шесть дней он уже прочно закреплен.
 отек мозга, который приводит к временному нарушению синап- тических связей между нейронами. У больных, подвергающихся лечению электрошоком, наступает полная амнезия на события, происшедшие в течение очень короткого периода перед воздействием; память о них никогда не возвращается, и это говорит о том, что электрошок нарушает только кратковременную память. Мак-Гоу (McGaugh) полагает, что при этом разрываются функциональные реверберирующие цепи. Это позволило бы объяснить результаты наших опытов, в которых электро
отек мозга, который приводит к временному нарушению синап- тических связей между нейронами. У больных, подвергающихся лечению электрошоком, наступает полная амнезия на события, происшедшие в течение очень короткого периода перед воздействием; память о них никогда не возвращается, и это говорит о том, что электрошок нарушает только кратковременную память. Мак-Гоу (McGaugh) полагает, что при этом разрываются функциональные реверберирующие цепи. Это позволило бы объяснить результаты наших опытов, в которых электрошок полностью уничтожал потенциалы, выработанные у кошек за один день при сравнительно небольшом числе предъявлений парных стимулов (около 200). Эти результаты были истолкованы как подтверждение того, что у животных создавались следы памяти в виде реверберирующих нейронных цепей. Сочетавшиеся сенсорные стимулы, разделенные промежутком в 200—400 мс, запускали циклическую активность соответствующей периодичности. Такой циклический поток импульсов состоит из бинарных элементов, но суммарный результат оказывается аналоговым—это вызванный потенциал, продукт сум- мации постсинаптических потенциалов (рис. 54).
Согласно гипотезе реверберирующих цепей, кратковременная память связана с изменениями только в мембранах нейронов. Поток импульсов, достигая высших уровней центральной нервной системы, запускает волну потенциалов действия типа «всё или ничего» в соответствующем замкнутом нейронном пути, но это ритмическое изменение потенциала не выходит за пределы мембраны и ее непосредственной близости — во всяком случае, так мы думаем теперь.
Долговременное (перманентное) хранение информации.
Первая возможность: реорганизация синапсов
Большинство авторов согласно в том, что длительное сохранение следов памяти требует структурных изменений в центральных нейронах. Для объяснения длительного хранения информации предложены две хорошо известные гипотезы. Одна из них связывает долговременную память с усилением синаптических связей между нейронами, а вторая — с внутриклеточным хранением следов памяти.
В 1955 г. Сентаготаи (Szentagothai) описал различия в величине поверхностей синапсов в спинном мозгу в зависимости от их использования. Чтобы сделать суть этого открытия более понятной, опишем некоторые особенности синапсов — функциональных элементов, обеспечивающих передачу импульсов с одного нейрона на другой.
Строение синапсов. Аксоны одного нейрона образуют контакты с телом или дендритом следующего нейрона посредством концевых вздутий. Как эти вздутия, содержащие пузырьки, так и протоплазма следующей клетки окружены мембраной толщиной около 50 А. Как показала электронная микроскопия, пре- сииаптическая мембрана отделена от постсинаптической мембраны щелью шириной 200 А. В пресинаптических окончаниях заключено особое вещество — медиатор. Каждый приходящий импульс вызывает освобождение медиатора и переход его через синаптическую щель. С помощью электронного микроско
па показано, что медиатор хранится в пузырьках, часть которых открывается в синаптическую щель.
ПСП — аналоговый сигнал. Если к синапсу не приходят импульсы, регистрация при помощи электродов, введенных в пост- синаптический нейрон около мембраны, выявляет состояние покоя. При стимуляции пресинаптического волокна между двумя сторонами мембраны регистрируется разность потенциалов, которую называют постсинаптическим потенциалом (ПСП). Это локальный ответ, и он отличается от потенциала действия тем, что не следует закону «всё или ничего» и ограничен постсинаптическим участком мембраны. Он представляет собой аналоговый сигнал, способный генерировать в примыкающей мембране новую серию потенциалов действия (рис. 55).
В некоторых клетках медиаторы вызывают только деполя- ризационные ПСП, т. е. снижают потенциал покоя. В других клетках другие медиаторы вызывают гиперполяризационные ПСП — повышают потенциал покоя. Деполяризационные потенциалы — возбуждающие, а гиперполяризационные —тормозные. Как уже говорилось, ПСП представляет собой местный феномен, он не распространяется и служит только для запуска потенциалов действия, а сам ПСП генерируется под влиянием приходящего сигнала, т. е. потенциала действия преси- наптической клетки. Таким образом, в этой точке передача нервных импульсов происходит путем превращения бинарных сигналов в аналоговые с последующим превращением снова в бинарные сигналы — импульсы.
Медиаторы. Как полагают, в пузырьках синаптических концевых вздутий различных нейронов образуются и хранятся несколько видов веществ-медиаторов. Они служат или для передачи импульсов, или для ее торможения. Ацетилхолин служит возбуждающим медиатором; введя его, можно искусственно вызвать деполяризацию. Тормозным медиатором считают гамма-аминомасляную кислоту (ГАМК).
Таким образом, потенциал действия пресинаптического нейрона доходит только до концевого вздутия аксона, где он вызывает освобождение медиатора. Этот последний переходит через синаптическую щель на мембрану постсинаптического нейрона и изменяет ее ионную проницаемость. В зависимости от количества медиатора генерируется ПСП большей или меньшей величины. Если ПСП деполяризационный и достаточно большой, он порождает периодические потенциалы действия, которые передаются по отходящему от этой второй клетки аксону (рис. 56).
Два вида мембраны. В соответствии с этой теорией каждый нейрон должен обладать мембраной двух видов: одной напротив концевых вздутий и второй, покрывающей всю остальную клетку. Такие же два вида мембраны имеются у рецепторов,
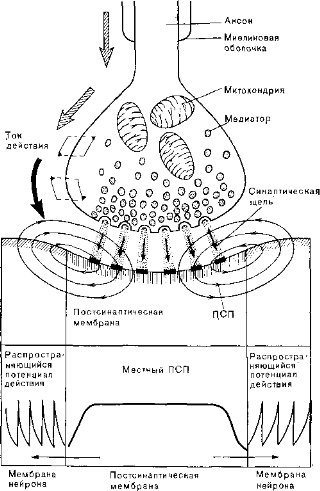
Рис. 55. Синаптическая передача импульсов. Область постсинаптических изменений, вызываемых медиатором, указана вертикальной штриховкой (здесь возникает местный ПСП).
Показано, что мембрана волокна в рецепторе способна генерировать локальный аналоговый сигнал, а в волокне вне рецептора регистрируются распространяющиеся сигналы типа «всё или ничего». Таким образом, ПСП — это генераторный потенциал, сходный с рецепторным потенциалом в местах воздействия сенсорных стимулов.
Синаптическая задержка. Время между приходом пресинап- тического импульса к концевому вздутию и генерацией ПСП, называемое синаптической задержкой, измерялось при помощи микроэлектродов. Установлено, что у разных животных оно варьирует от 0,3 до 3 мс. Эта задержка складывается из отрезков времени, необходимых 1) для высвобождения медиатора, 2) для его диффузии к постсинаптической мембране и 3) для генерации ПСП.
Рост синаптических структур. Рассмотрим теперь возможную роль синапсов в хранении следов памяти. Как полагают, абсолютное число синапсов у одного нейрона и величина концевых вздутий зависят от использования пути, к которому они
Хим. Анал, С. Вин С.
|
|
0 |
|
Потенциал |
? |
|
действия |
э |
|
|
с |
|
Мембрана |
Ч) 3 |
|
неирока |
с |
|
|
|
|
|
|
ф |
|
|
|
|
|
Местный |
Потенциал |
|
|
ПСП |
действия |
|
|
|
|
|
|
|
|
О |
|
Постен • к )птиче- |
Мембрана |
3 |
|
мембранг |
нейрона |
X. |
|
Местны й ПСП |
Потенциал действия |
|
|
Постси . |
Мембрана |
|
|
|
нейрона |
|
|
мембран; |
|
|
|
Местны й ПСП |
|
|
Постси- |
Мембрана |
|
с«ая |
нейронд |
|
мембране |
|
Направление передачи импульса
Рис. 56. Химическая передача, делающая возможным преобразование бинарных сигналов в аналоговые при распространении нервных импульсов по цепи нейронов.
принадлежат. При более интенсивном использовании эти параметры имеют тенденцию возрастать; и напротив, они уменьшаются, если проводящий путь бездействует. Это говорит о хорошей адаптационной способности аппарата, передающего импульсы. Изменения величины синапсов продемонстрированы гистологическими методами.
Более длительные ПСП. Экклс (Eccles) показал, что в высших нервных образованиях (головном мозгу) способность синапсов к адаптации выражена сильнее, чем на низших уровнях, например в спинном мозгу или в стволе мозга. Амплитуда и длительность ПСП в головном мозгу примерно в десять раз больше, чем в спинном, где ПСП, как правило, слабые и кратковременные (рис. 57). Поэтому Экклс полагает, что интенсивное использование синапсов в мозгу приводит к увеличению не только их поверхностей, но также и количества медиаторов, которые вызывают более длительный ПСП.
Посттетаническая потенциация. Адаптационная способность синапсов выявляется в опытах с созданием посттетанической потенциации. Электрическое раздражение током высокой частоты (тетанизация) на протяжении нескольких минут ведет к облегчению передачи импульсов, т. е. к понижению порога постсннаптической мембраны. В постсинаптической клетке в течение нескольких минут после тетанизации потенциал действия будет возникать в ответ даже на стимулы, бывшие ранее подпороговыми. Нейрон как бы «помнит» высокочастотную стимуляцию, которая прекратилась за несколько минут до этого. Возможно, что подобный механизм участвует и в синаптическом процессе образования следов памяти.
Резюме: три вида изменений в синапсах. Итак, известны три разных свойства синапсов, которые могли бы служить физио-
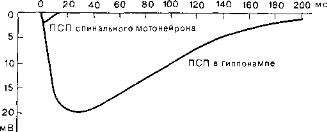
Рис. 57. График Экклса, показывающий, что в головном мозгу ПСП длятся дольше, чем в спинном.
логической основой длительного сохранения следов памяти: увеличение синаптических поверхностей, большая продолжительность ПСП и снижение порога, сходное с постсинаптической потенциацией. Все эти три изменения могли бы участвовать в создании свойства, характерного для нейронных путей в головном мозгу, которое состоит в том, что эти пути становятся более чувствительными по мере их использования, когда они, так сказать, «проторяются».
Ускоренный синтез белка. Никакое увеличение объема концевых вздутий и усиление выработки медиатора нельзя себе представить без ускорения сложных молекулярных процессов внутри клетки. Самый главный из них, несомненно, состоит в синтезе белков, управляемом РНК- В последние годы опубликованы данные, которые показывают, что в росте синапсов при их усиленном функционировании может играть роль образование полипептидпых цепей. Следует подчеркнуть, что это ускорение синтеза белка не идентично хранению информации внутри клетки, о которой будет сказано ниже. В этом случае не
возникают новые белки с иной последовательностью аминокислот, а только ускоряется синтез белков в концевых вздутиях, что ведет к количественным изменениям в синаптическом аппарате и усиленной выработке медиатора.