
Профаза I
Как уже отмечалось (см. главу 1), ооциты человека вступают в мейоз на ранних стадиях эмбрионального развития. Несмотря на труд- нодоступность материала, неоднократно предпринимались попытки изучить поведение хромосом в профазе I с помощью традиционных цитогенетических методов или путем анализа тотальных СК [246, 556, 586, 592, 686, 803, 821]. Молекулярно-цитогенетические и иммуноцитохимические методы предоставили новые возможности для анализа процессов конъюгации и рекомбинации в оогенезе, однако работы такого рода единичны [225, 296, 297].
К настоящему времени на материале медицинских абортусов 6-24 недель развития установлена хронология профазы I. Так, ооциты на стадии лептотены — зиготены появляются на 9-й неделе и сохраняются в яичниках плодов до 24-й недели беременности, первые ооциты на стадиях пахитены — диплотены зарегистрированы одновременно с дикти- отеной в 13-14 недель [357]. Известно также, что у новорожденных все ооциты в яичнике находятся на стадии диктиотены. Аналогичные данные
Таблица 4.2. Материал, доступный для цитогенетического анализа оогенезау человека
Незрелые ооциты
Зрелые ооциты
Биопсия эмбриона

развивающихся яичниках у плодов с врожденными пороками развития и хромосомными болезнями нами были предприняты специальные исследования [86]. Хронология профазы I была изучена на хромосомных препаратах, полученных из аутопсийного материала яичников от 58 плодов женского пола сроком развития 19-26 недель беременности. В 12 случаях материал был получен от спонтанных абортусов, в 31 случае — после прерывания беременности по социальным показаниям и в 15 — в связи с наличием хромосомных болезней (n = 7) или врожденными пороками развития (n = 8). По результатам патоморфологического исследования, все индуцированные абортусы, полученные в результате прерывания беременности по социальным показаниям, соответствовали сроку беременности и не имели пороков развития. Поэтому мы сочли возможным использовать их в качестве условно нормальной, контрольной группы. Все плоды из группы спонтанных абортусов имели признаки внутриутробной задержки развития, сопровождавшиеся плацентарной недостаточностью. Врожденные пороки развития были представлены пороками нервной и сердечно-сосудистой систем, а хромосомные — трисомией 21 (n = 4) и трисомией 18 (n = 1).
Сравнительный анализ распределения ооцитов по стадиям профазы показал, что в случаях остановки или замедления темпов внутриутробного развития при отсутствии аномалий морфогенеза отмечается задержка мейоза на стадии лептотены. Формирование множественных пороков развития, включая нервную и сердечно-сосудистую системы сопровождается ускорением темпа мейоза, которое выражается в снижении численности ооцитов начальных стадий профазы и резком увеличении числа ооцитов на стадиях пахитены — диплотены. Влияние трисомии 21 и 18 на динамику профазы проявляется двояким образом. Часть ооцитов в ускоренном темпе преодолевает начальные стадии профазы, достигая стадии диплотены, тогда как остальные проходят стадии прелептотены — лептотены в замедленном темпе или претерпевают остановку на стадии зиготены. При этом в большинстве ооцитов у плодов с трисомией триада хромосом 21 представлена унивалентами, что, по-видимому, является одним из признаков их неспособности к завершению профазы мейоза.
К сожалению, следует признать, что сведения об изменении хронологии начальных стадий оогенеза недостаточны для понимания механизмов действия тех или иных факторов на профазу первого мейоти- ческого деления. Однако полученные нами результаты иллюстрируют
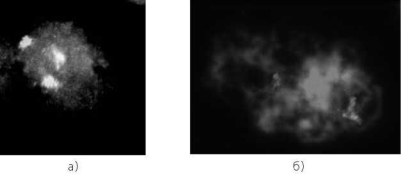
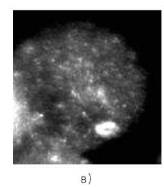
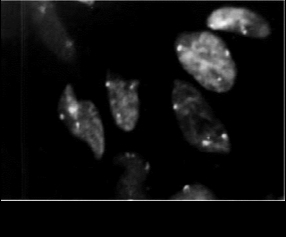
Рис. 4.7. Мейоциты из яичника от медицинского абортуса (кариотип 47,ХХ, +21) сроком развития 21/22 недели. FISH с цельнохромосомным ДНК-зондом к хромосоме 21. Варианты конъюгации хромосом 21: а — полный асинапсис хромосом триады, б — бивалент + унивалент, в — тривалент
многообразие причин, приводящих к изменению динамики мейотических преобразований в эмбриональный и плодный периоды развития человека.
Особый интерес представляют результаты анализа профазных хромосом у плодов с несбалансированным кариотипом (рис. 4.7). Так, при трисомии 21 и 18 наблюдается как полный асинапсис хромосом триады, так и конъюгация гомологов, которая осуществляется двояким образом — бивалент + унивалент или тривалент [225, 297]. СК в трива- ленте формируется либо по всей длине триады хромосом 21, либо частично, с образованием «шпилек» [882].
Как известно, в ходе эмбрионального развития наблюдается массовая гибель ооцитов (см. главу 1). Предполагается, что одной из причин такой гибели является аномальная конъюгация хромосом [640, 802]. Повышенная частота атретичных ооцитов у анеуплоидных плодов [225] и полное отсутствие мейоцитов у плода 69,ХХХ (рис. 4.8) [88] согласуются с этой гипотезой. Однако механизмы, регулирующие вступление ооцитов в апоптоз, до сих пор не выяснены [647]. Таким образом, профаза мейоза у эмбрионов женского пола с нормальным и гетероплоидным кариотипом является уникальной естественной моделью для изучения конъюгации хромосом, антенатальной селекции ооцитов и анализа контролирующих механизмов запрограммированной клеточной гибели — апоптоза.
Как уже отмечалось (см. главу 1), ооциты человека вступают в мейоз на ранних стадиях эмбрионального развития. Несмотря на труд- нодоступность материала, неоднократно предпринимались попытки изучить поведение хромосом в профазе I с помощью традиционных цитогенетических методов или путем анализа тотальных СК [246, 556, 586, 592, 686, 803, 821]. Молекулярно-цитогенетические и иммуноцитохимические методы предоставили новые возможности для анализа процессов конъюгации и рекомбинации в оогенезе, однако работы такого рода единичны [225, 296, 297].
К настоящему времени на материале медицинских абортусов 6-24 недель развития установлена хронология профазы I. Так, ооциты на стадии лептотены — зиготены появляются на 9-й неделе и сохраняются в яичниках плодов до 24-й недели беременности, первые ооциты на стадиях пахитены — диплотены зарегистрированы одновременно с дикти- отеной в 13-14 недель [357]. Известно также, что у новорожденных все ооциты в яичнике находятся на стадии диктиотены. Аналогичные данные
Таблица 4.2. Материал, доступный для цитогенетического анализа оогенезау человека
- Оогонии и ооциты в профазе первого деления мейоза
- эмбрионы 9-26-й недель развития
- При использовании методов ЭКО*:
Незрелые ооциты
- стадия зародышевого пузырька (профаза первого деления мейоза, поздняя диплотена
- разрушение зародышевого пузырька (диакинез)
- метафаза первого деления мейоза (MI)
Зрелые ооциты
- метафаза второго деления мейоза (MII) + первое полярное тельце (ППТ)
- изолированное ППТ (биопсия, выполненная для генетической диагностики) Ооциты, используемые для оплодотворения in vitro
- неоплодотворившиеся ооциты в MII
- оплодотворившиеся ооциты
- нормальные зиготы (с женским и мужским пронуклеусами) + второе полярное тельце (ВПТ)
- изолированное ВПТ (биопсия, выполненная для генетической диагностики)
- аномальное оплодотворение (полиплоидная зигота)
- незавершенное оплодотворение: хромосомы в MII + преждевременно конденсированные хромосомы отцовского пронуклеуса, хроматиновые массы и т. д.
- ооциты, активированные к партеногенетическому развитию Дробящиеся эмбрионы
- нормальное развитие
- аномальное развитие (остановка дробления, фрагментация)
Биопсия эмбриона
- изолированные бластомеры от эмбриона на стадии дробления
- биопсия трофэктодермы на стадии бластоцисты
развивающихся яичниках у плодов с врожденными пороками развития и хромосомными болезнями нами были предприняты специальные исследования [86]. Хронология профазы I была изучена на хромосомных препаратах, полученных из аутопсийного материала яичников от 58 плодов женского пола сроком развития 19-26 недель беременности. В 12 случаях материал был получен от спонтанных абортусов, в 31 случае — после прерывания беременности по социальным показаниям и в 15 — в связи с наличием хромосомных болезней (n = 7) или врожденными пороками развития (n = 8). По результатам патоморфологического исследования, все индуцированные абортусы, полученные в результате прерывания беременности по социальным показаниям, соответствовали сроку беременности и не имели пороков развития. Поэтому мы сочли возможным использовать их в качестве условно нормальной, контрольной группы. Все плоды из группы спонтанных абортусов имели признаки внутриутробной задержки развития, сопровождавшиеся плацентарной недостаточностью. Врожденные пороки развития были представлены пороками нервной и сердечно-сосудистой систем, а хромосомные — трисомией 21 (n = 4) и трисомией 18 (n = 1).
Сравнительный анализ распределения ооцитов по стадиям профазы показал, что в случаях остановки или замедления темпов внутриутробного развития при отсутствии аномалий морфогенеза отмечается задержка мейоза на стадии лептотены. Формирование множественных пороков развития, включая нервную и сердечно-сосудистую системы сопровождается ускорением темпа мейоза, которое выражается в снижении численности ооцитов начальных стадий профазы и резком увеличении числа ооцитов на стадиях пахитены — диплотены. Влияние трисомии 21 и 18 на динамику профазы проявляется двояким образом. Часть ооцитов в ускоренном темпе преодолевает начальные стадии профазы, достигая стадии диплотены, тогда как остальные проходят стадии прелептотены — лептотены в замедленном темпе или претерпевают остановку на стадии зиготены. При этом в большинстве ооцитов у плодов с трисомией триада хромосом 21 представлена унивалентами, что, по-видимому, является одним из признаков их неспособности к завершению профазы мейоза.
К сожалению, следует признать, что сведения об изменении хронологии начальных стадий оогенеза недостаточны для понимания механизмов действия тех или иных факторов на профазу первого мейоти- ческого деления. Однако полученные нами результаты иллюстрируют
Рис. 4.7. Мейоциты из яичника от медицинского абортуса (кариотип 47,ХХ, +21) сроком развития 21/22 недели. FISH с цельнохромосомным ДНК-зондом к хромосоме 21. Варианты конъюгации хромосом 21: а — полный асинапсис хромосом триады, б — бивалент + унивалент, в — тривалент
многообразие причин, приводящих к изменению динамики мейотических преобразований в эмбриональный и плодный периоды развития человека.
Особый интерес представляют результаты анализа профазных хромосом у плодов с несбалансированным кариотипом (рис. 4.7). Так, при трисомии 21 и 18 наблюдается как полный асинапсис хромосом триады, так и конъюгация гомологов, которая осуществляется двояким образом — бивалент + унивалент или тривалент [225, 297]. СК в трива- ленте формируется либо по всей длине триады хромосом 21, либо частично, с образованием «шпилек» [882].
Как известно, в ходе эмбрионального развития наблюдается массовая гибель ооцитов (см. главу 1). Предполагается, что одной из причин такой гибели является аномальная конъюгация хромосом [640, 802]. Повышенная частота атретичных ооцитов у анеуплоидных плодов [225] и полное отсутствие мейоцитов у плода 69,ХХХ (рис. 4.8) [88] согласуются с этой гипотезой. Однако механизмы, регулирующие вступление ооцитов в апоптоз, до сих пор не выяснены [647]. Таким образом, профаза мейоза у эмбрионов женского пола с нормальным и гетероплоидным кариотипом является уникальной естественной моделью для изучения конъюгации хромосом, антенатальной селекции ооцитов и анализа контролирующих механизмов запрограммированной клеточной гибели — апоптоза.