
Особый интерес представляет метилирование хромосом при анеуплоидии. Паттерн метилирования триады хромосом был изучен нами у двух зародышей с разными трисомиями.
В случае трисомии 16 показанием к прерыванию беременности в срок 6/7 недель явилась анэмбриония, т. е. отсутствие эмбриона в плодном пузыре. Благодаря гетероморфизму гетерохроматиновых блоков хромосомы 16 удалось установить, что нерасхождение произошло в оогенезе, в первом делении мейоза. После ник-трансляции на препаратах из клеток цитотрофобласта, предобработанных MspI, сигналы были выявлены в районе 16q11 во всех трех гомологах, а после
*
предобработки HpaII — лишь у двух гомологов из триады (рис. 10.20) . При этом отсутствие ник-трансляционного сигнала зарегистрировано в гетерохроматиновом блоке хромосомы 16 материнского происхождения во всех клетках. У эмбрионов с нормальным кариотипом такого
же срока гетерохроматин хромосомы 16 обоих гомологов характеризовался наличием сигнала. Известно, что трисомия 16 является наиболее частой хромосомной аномалией у ранних спонтанных абортусов (31 % от всех трисомий), особенно при анэмбрионии. При развивающейся беременности она крайне редка и ограничена исключительно цито- трофобластом. Возможно, в первом триместре дисбаланс, обусловленный дополнительной хромосомой 16, частично компенсируется за счет метилирования и инактивации, возможно, «ранних генов», локализованных в прицентромерном гетерохроматине. Этот механизм, однако, оказывается неэффективным на более поздних стадиях. Данное предположение требует, несомненно, более детальных исследований.
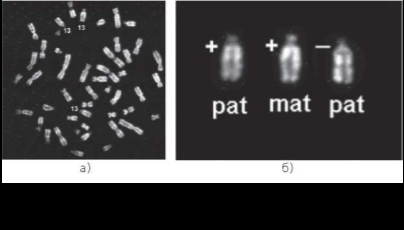
Характер метилирования был изучен также в ФГА-стимулированных лимфоцитах пуповинной крови плода 23/24 недель развития с кариотипом 47,ХХ,+13. По гетероморфизму коротких плеч хромосом 13 было установлено отцовское происхождение дополнительной хромосомы при нерасхождении в первом делении мейоза. При сравнении характера метилирования гомологов хромосомы 13 выявлено различие в районе 13q11, являющемся R-сегментом. Так, на дополнительной хромосоме отцовского происхождения в этом участке хромосомы после обработки HpaII сигнал отсутствовал, т. е. этот сегмент был гиперметилирован, тогда как после обработки MspI было характерно включение меченых нуклеотидов в тождественный район всех трех хромосом (рис. 10.21).
Таким образом, в обоих случаях трисомии было зарегистрировано изменение статуса метилирования отдельных локусов на дополнительных хромосомах, что косвенно указывало на их инактивацию. Возможно, таким способом осуществляется реализация некоего «компенсаторного механизма». Безусловно, гипотеза об избирательной компенсации дозы генов на уровне хромосомных сегментов в случае наличия дополнительных хромосом в кариотипе основана пока на единичных фактах. Однако их накопление и детальный анализ, возможно, позволят выяснить механизмы, действующие в несбалансированных кариотипах.
В случае трисомии 16 показанием к прерыванию беременности в срок 6/7 недель явилась анэмбриония, т. е. отсутствие эмбриона в плодном пузыре. Благодаря гетероморфизму гетерохроматиновых блоков хромосомы 16 удалось установить, что нерасхождение произошло в оогенезе, в первом делении мейоза. После ник-трансляции на препаратах из клеток цитотрофобласта, предобработанных MspI, сигналы были выявлены в районе 16q11 во всех трех гомологах, а после
*
предобработки HpaII — лишь у двух гомологов из триады (рис. 10.20) . При этом отсутствие ник-трансляционного сигнала зарегистрировано в гетерохроматиновом блоке хромосомы 16 материнского происхождения во всех клетках. У эмбрионов с нормальным кариотипом такого
же срока гетерохроматин хромосомы 16 обоих гомологов характеризовался наличием сигнала. Известно, что трисомия 16 является наиболее частой хромосомной аномалией у ранних спонтанных абортусов (31 % от всех трисомий), особенно при анэмбрионии. При развивающейся беременности она крайне редка и ограничена исключительно цито- трофобластом. Возможно, в первом триместре дисбаланс, обусловленный дополнительной хромосомой 16, частично компенсируется за счет метилирования и инактивации, возможно, «ранних генов», локализованных в прицентромерном гетерохроматине. Этот механизм, однако, оказывается неэффективным на более поздних стадиях. Данное предположение требует, несомненно, более детальных исследований.
Характер метилирования был изучен также в ФГА-стимулированных лимфоцитах пуповинной крови плода 23/24 недель развития с кариотипом 47,ХХ,+13. По гетероморфизму коротких плеч хромосом 13 было установлено отцовское происхождение дополнительной хромосомы при нерасхождении в первом делении мейоза. При сравнении характера метилирования гомологов хромосомы 13 выявлено различие в районе 13q11, являющемся R-сегментом. Так, на дополнительной хромосоме отцовского происхождения в этом участке хромосомы после обработки HpaII сигнал отсутствовал, т. е. этот сегмент был гиперметилирован, тогда как после обработки MspI было характерно включение меченых нуклеотидов в тождественный район всех трех хромосом (рис. 10.21).
Таким образом, в обоих случаях трисомии было зарегистрировано изменение статуса метилирования отдельных локусов на дополнительных хромосомах, что косвенно указывало на их инактивацию. Возможно, таким способом осуществляется реализация некоего «компенсаторного механизма». Безусловно, гипотеза об избирательной компенсации дозы генов на уровне хромосомных сегментов в случае наличия дополнительных хромосом в кариотипе основана пока на единичных фактах. Однако их накопление и детальный анализ, возможно, позволят выяснить механизмы, действующие в несбалансированных кариотипах.