
При исследовании ЯОР на «прямых» препаратах метафазных хромосом из клеток экстраэмбриональных (цитотрофобласт) и эмбриональных тканей (мозг, кишечный эпителий, легкое, печень, почка, роговица глаза) медицинских абортусов с нормальным кариотипом оказалось, что среднее число Ag+^ОР хромосом на метафазную пластинку варьирует от 6,1 ± 0,46 до 8,3 ± 0,68 [146].
Средняя суммарная активность ЯОР хромосом групп D и G, отражающая интенсивность экспрессии локализованных в них р-генов, варьировала в широких пределах — от 17,8 ± 1,64 до 34,2 ± 2,18 баллов [146]. При этом 42 % общей вариабельности активности р-генов составляли индивидуальные различия. Межиндивидуальные различия суммарной активности, определяемые комбинаторикой родительских хромосом, ранее отмечались и другими исследователями [107, 122, 180].
Особый интерес представляло сопоставление активности ЯОР хромосом цитотрофобласта с другими эмбриональными тканями. Известно, что гены рРНК являются ключевым компонентом путей клеточной сигнализации, контролирующих рост и пролиферацию клеток [652]. Учитывая активный рост ворсин хориона при формировании хорион-аллантоидной плаценты и интенсивную пролиферацию клеток цитотрофобласта [734], а также увеличение активности ЯОР в быстро делящихся клетках [140, 238, 631], логично предполагать увеличение транскрипционной активности р-генов в клетках цитотрофобласта по сравнению с другими эмбриональными тканями. Результаты исследований, однако, свидетельствуют об отсутствии явных отличий активности ЯОР в клетках цитотрофобласта по сравнению с другими эмбриональными тканями [122, 146].
Следует отметить, что имеющиеся сведения о влиянии типа ткани на функционирование р-генов противоречивы. Согласно одним авторам, функциональный статус ЯОР в эмбриональных клетках не зависит от тканевой принадлежности и соответствует таковому в бласт- трансформированных лимфоцитах периферической крови взрослых [117, 146, 180]. Результаты других исследований, напротив, свидетельствуют о значительной вариабельности экспрессии р-генов в разных тканях одного индивидуума [278, 600, 629]. Причины наблюдаемых различий не совсем ясны. Не исключено, однако, что они обусловлены методическими особенностями приготовления препаратов, в частности, использованием в большинстве исследований клеточных культур. Между тем, условия in vitro, возможно через механизм метилирования ДНК [22, 422], могут существенно влиять на экспрессию рибосомных генов [261].
Таким образом, в клетках различных тканей у эмбрионов человека 6-14 недель развития как по числу Ag+^ОР хромосом, так и по суммарной активности локализованных в них р-генов, наблюдается межхромосомный, межклеточный и межиндивидуальный полиморфизм, не отличающийся по диапазону от функционального полиморфизма ЯОР в постнатальный период онтогенеза.
Как показывают наши исследования, суммарная транскрипционная активность ЯОР у эмбрионов человека 6-14 недель развития возрастает по мере развития плода (рис. 10.3), тогда как зависимость числа Ag+^ЯОР хромосом на клетку от срока беременности прослеживается не столь отчетливо.
Увеличение транскрипционной активности рибосомных генных кластеров носит хромосом-специфичный характер. Так, р-гены, локализованные на хромосомах группы G, экспрессируются интенсивнее, чем на хромосомах группы D. Так как число Ag+^ОР хромосом не зависит от стадии эмбриогенеза, увеличение уровня суммарной активности ЯОР можно объяснить большей функциональной активностью р-генных кластеров хромосом 21 и 22, по сравнению с хромосомами 13, 14, 15 [146].
Предполагается, что наследуемая программа экспрессии рибосомных генов реализуется за счет «резервных» молчащих р-генов [103], которые, возможно, могут активироваться в связи с необходимостью интенсивного белкового синтеза в процессе эмбриогенеза. При этом число резервных р-генов варьирует на разных акроцентрических хромосомах, и их активация может происходить неравномерно. В частности, хромосома 21, по сравнению с другими ЯО-хромосомами, содержит наибольшее число молчащих копий рибосомных генов, а хромосома 15 имеет пониженную активность и является наиболее низкоко- пийной по содержанию р-генов [103].
Таким образом, основной причиной полиморфизма ЯОР хромосом эмбрионов человека является комбинаторика родительских ЯОР хромосом, определяющая индивидуальные различия. Нельзя, однако, исключить и наличия реальных молекулярных механизмов репрес- сии/дерепрессии р-генов, регулирующих модуляции их активности в процессе онтогенеза. Согласно последним данным [450], регуляция активности ЯОР районов может осуществляться с помощью одного из трех механизмов: элиминации части рДНК, ее метилирования, эффекта положения гетерохроматиновых районов. Какой из этих механизмов модуляции активности ЯОР превалирует в эмбриогенезе человека остается неизвестным и требует специального изучения.
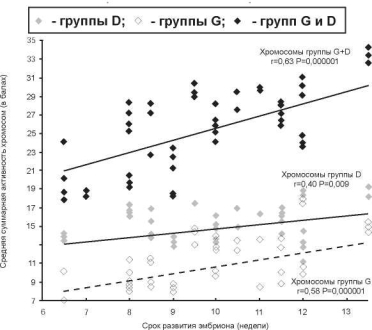
Рис. 10.3. Динамика активности ЯОР хромосом в цитотрофобластеу эмбрионов человека с нормальным кариотипом в 6—14 недель развития
В 6-14 недель, во время активного органогенеза, интенсивность транскрипции р-генов возрастает. Можно предполагать, что этот феномен обусловлен увеличением числа работающих копий генов и/или скоростью транскрипции, а не увеличением числа аргентофильных ЯО-хромосом. Двухступенчатая модель активации р-генов (см. выше) хорошо согласуется с этим предположением. Согласно данной модели, первичная активация транскрипции происходит в ядрышковых организаторах с наибольшим числом рибосомных цистронов; активация других ЯОР начинается только после завершения транскрипции предыдущего. Другой причиной полиморфизма ЯОР является срок эмбрионального развития, который в большей степени сказывается на хромосомах группы G, нежели группы D. Эти результаты указывают на различную функциональную значимость хромосом G и D в эмбриогенезе человека.
- Полиморфизм ЯОР у плодов 20-24 недель развития
Для характеристики состояния ЯОР в эмбриогенезе человека во второй половине беременности были проведены исследования метафазных хромосом из ФГА-стимулированных лимфоцитов пуповинной крови, полученной при кордоцентезе, от 43 плодов с нормальным кариотипом (2n = 46) в срок 20-24 недель беременности. В 22 случаях пренатальное кариотипирование было выполнено в связи с высоким риском хромосомных болезней у плода: возраст матери 39 лет и старше, наличие хромосомной болезни или множественных пороков развития у предыдущего ребенка (группа 1). В 8 случаях — для выработки тактики родоразрешения в связи с наличием у плода пороков развития, выявленных при ультразвуковом исследовании (группа 2), и в 13 — с целью пренатальной диагностики генных болезней (группа 3).
У всех плодов наблюдалась межклеточная и межиндивидуальная вариабельность числа Ag+^ромосом и активности их ЯОР. Ранжирование случаев по возрастанию суммарной активности ЯОР хромосом позволили сформировать группы сравнения, которые, что оказалось неожиданным, соответствовали показаниям для проведения кордо- центеза. Интересно, что в группах плодов, имевших повышенный риск хромосомных болезней (группа 1), или пороки развития, не совместимые с живорождением и постнатальным развитием (группа 2), среднее число Ag+^ромосом на клетку оказалось выше, чем у плодов с популяционным риском хромосомных болезней (группа 3). В группах 1 и 2 были сильнее выражены межклеточная и межиндивидуальная вариабельность суммарной активности ЯОР по сравнению с группой контроля (табл. 10.1). Сравнение полигонов частот ЯОР показало, что распределения суммарной активности ЯОР между тремя группами различны (рис. 10.4), однако они хорошо соответствуют представлениям о пределах варьирования активности ЯОР хромосом в популяции здоровых индивидов с нормальным кариотипом [101, 107]. Так, в контрольной группе суммарная активность ЯОР фактически не выходила за пределы таковой для взрослой популяции и составляла 18-26 баллов (табл. 10.1).
У плодов с высоким риском анеуплоидии и с пороками развития отмечена значительная вариабельность активности ЯОР, а также ее смещение в сторону увеличения (19-38 баллов) (табл. 10.1, рис. 10.4). Эти
Таблица 10.1. Полиморфизм числа А?+-ЯОР хромосом и суммарной активности ЯОР в ФГА-стимулированных лимфоцитах пуповинной крови у плодов 20—24 недель развития (собственные данные)
|
Группы плодов |
Число плодов |
Среднее число Ag+^ОР хромосом на клетку |
Пределы вариабельности суммарной активности ЯОР (в баллах) |
|
|
1 |
Высокий риск хромосомной болезни у плода |
22 |
8,37 ±0,104* |
19,3 ±2,39 — 37,2 ±1,83 |
|
2 |
Грубые пороки развития у плода |
8 |
8,67 ±0,131* |
19,0 ± 2,76 — 38,3 ± 1,07 |
|
3 |
Контрольная группа (высокий риск моногенных болезней у плода) |
13 |
7,25 ±0,081 |
16,9 ± 1,09 — 29,5 ± 0,70 |
*Р lt; 0,05.
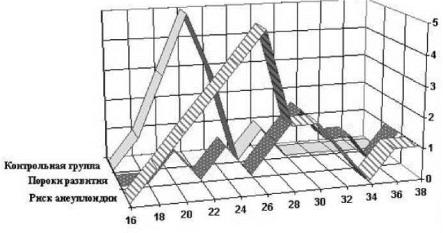
Рис. 10.4. Распределение суммарной активности ЯОР хромосом в ФГА-стимулиро- ванных лимфоцитах из пуповинной крови плодов человека с нормальным кариотипом в 20—24 недели развития. По оси абсцисс — средняя активность ЯОР на клетку (в баллах), по оси ординат — число плодов
наблюдения хорошо согласуются с результатами сравнительного исследования полиморфизма ЯОР хромосом у спонтанных абортусов 9-16 недель беременности [117]. Хотя сравнение данных разных авторов затруднено вследствие различий критериев визуальной оценки активности ЯОР, нельзя не обратить внимания на общую тенденцию к смещению в сторону увеличения суммарной активности ЯОР у плодов с пониженной жизнеспособностью. Логично предположить, что процессы, связанные с нарушением жизнеспособности плода и приводящие к его внутриутробной гибели и/или спонтанному прерыванию беременности, существенно влияют и на функциональную активность р-генов.
Угрозой прерывания беременности, по-видимому, можно объяснить и смещение распределения суммарной активности ЯОР у плодов группы цитогенетического скрининга (группа 1). Предполагается, что у спонтанных абортусов с нормальным кариотипом увеличение суммарной активности ЯОР в геноме может быть связано с адаптивными механизмами ответа системы рибосомных генов на неблагоприятные условия развития [117]. К сожалению, мы не располагаем сведениями о дальнейшем течении этих беременностей и их исходах. Нельзя, однако, исключить и другие причины увеличения суммарной активности ЯОР хромосом у плодов этой группы. В частности, у родителей детей с синдромом Дауна обнаружена высокая частота экстремальных вариантов ЯОР, в т. ч. двойных ЯОР [857]. Кроме того, некоторые авторы отмечают тенденцию к увеличению активности р-генов у женщин по мере увеличения репродуктивного возраста [117]. В любом случае, плоды именно таких родителей могут унаследовать хромосомы с повышенной экспрессией /-генов, что и определяет наличие у них высокого уровня суммарной активности ЯОР. Для проверки данного предположения необходимы сведения о функциональном статусе ЯОР родительских хромосом и изучение характера их наследования.
Таким образом, при исследовании функционального полиморфизма ЯОР у плодов человека во втором триместре беременности установлено, что пределы вариабельности суммарной активности р-генов у плодов, матери которых имели невысокий риск рождения ребенка с хромосомной патологией, соответствуют установленным у здоровых индивидов после рождения. В то же время, при наличии у плодов с нормальным кариотипом пороков развития, несовместимых с жизнью, экспрессия р-генов возрастает. Аналогичная, но менее однознач-
ная ситуация наблюдается и для плодов у беременных групп высокого риска рождения ребенка с хромосомной болезнью. Увеличение активности ЯОР хромосом в лимфоцитах плода, по-видимому, свидетельствует о менее благоприятном прогнозе для внутриутробного развития. Выявленные нами закономерности функциональной гетерогенности ЯОР у плодов в зависимости от факторов риска хромосомной патологии требуют дальнейших исследований.