
Сохранение паттерна метилирования ДНК, характерного для каждого типа клеток, важно для обеспечения их нормальной жизнедеятельности и функции. Вместе с тем, в настоящее время накапливается все больше фактов, что его нарушение может играть существенную роль в процессах трансформации нормальных клеток в опухолевые. Изменения статуса метилирования широко распространены при спорадических раковых заболеваниях человека, характеризующихся гипометилированием на уровне генома и локальным абберантным гиперметилированием CpG-островков [861]. При этом оба процесса, одновременно происходящие в разных районах генома, могут предшествовать озлокачествлению [334]. Важная роль в онкогенезе принадлежит спонтанному дезаминированию 5-метилцитозина в определенных районах генов, которое приводит к точковым мутациям — заменам С на Т (рис. 10.10). 24 % мутаций в гене опухолевого супрессора белка p53 составляют замены С^-Т в динуклеотидах CpG, что является одной из причин, детерминирующих образование солидных опухолей [658, 836]. Известно, что в гене p53 содержится около 290 CpG-динуклеотидов. При этом точковые замены в 23 CpG-динуклеотидах, расположенных в районе, кодирующем ДНК-связывающий домен, составляют треть от общего числа мутаций в этом гене [560]. Установлена опре
деленная зависимость между расположением «горячих точек» мутаций и локализацией опухоли. Так, в гене р53 мутационные изменения в кодонах 175, 248, 273 характерны только для опухолей молочной железы, яичника и для лейкемии [798], а в кодоне 157 — лишь для опухолей легкого у курящих пациентов [709].
Существует три гипотезы, объясняющие роль гипометилирования в индукции злокачественных образований — активация протоонкогенов, хромосомная нестабильность и активация латентных ретротранспазонов [836]. При этом уровень озлокачествления в определенной мере коррелирует со степенью метилирования геномной ДНК [776, 795].
Важную роль в опухолевой трансформации нормальных клеток может играть и процесс гиперметилирования, в особенности, если он затрагивает промоторные области генов онкосупрессоров (рис. 10.10). Ген ретинобластомы pRb был первым из множества генов, для которых была доказана взаимосвязь между гиперметилированием их промоторов и инактивацией транскрипции [435]. Такой же механизм регуляции характерен и для генов, контролирующих клеточный цикл, дифферен- цировку, апоптоз, репарацию ДНК, устойчивость к наркотическим веществам, детоксикацию и т. д. [108]. Нередко гиперметилирование промоторной области генов коррелирует с увеличением уровня актив-
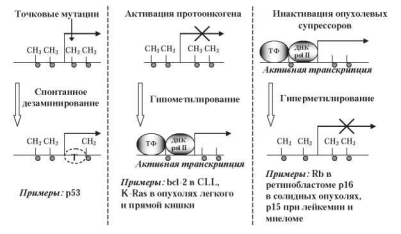
Рис. 10.10. Предполагаемое значение спонтанного дезаминирования 5-метилцитозина, гипометилирования и гиперметилирования в онкогенезе [792]
ности ДНК-метилтрансфераз, что ведет к стабильному выключению функций онкосупрессоров, а также генов, контролирующих клеточную пролиферацию или апоптоз [539].
Сегодня не вызывает сомнения, что изменение статуса метилирования может предшествовать началу опухолевой трансформации нормальных клеток при многих раковых заболеваниях. Считается, что дефекты метилирования, обусловленные действием различных эндогенных и экзогенных факторов в сочетании с предсуществующими мутациями онкогенов и онкосупрессоров, создают, согласно известной «двухударной гипотезе» [546], необходимые условия для бласт-трансформации и злокачественного роста [72]. Открытие этих механизмов послужило толчком для разработки многочисленных генетических тестов ранней диагностики, лечения и прогнозирования онкологических заболеваний на основе изучения статуса метилирования многих генов.
Важно отметить, что столь существенные в процессах онкогенеза гены, контролирующие клеточную пролиферацию, дифференцировку и апоптоз, являются, посути, «эмбриональными» генами, т. е. генами, ответственными за раннее эмбриональное развитие.