
Под аллергическим иммунным ответом следует понимать ТИ2-зави- симый гуморальный иммунный ответ, при котором образуются антитела IgE-класса. Аллергический иммунный ответ служит предпосылкой развития аллергических реакций (при условии повторного поступления аллергена).
ТН2-зависимый иммунный ответ как основа аллергии
Т-клеточный контроль аллергического ответа характеризуется доминирующим влиянием Th2-клеток на его развитие (рис. 4.26). При развитии первичного ответа на аллергены факторами, благоприятствующими формированию ^2-ответа, служат условия микроокружения дендритных клеток, поглощающих аллерген в слизистых оболочках. Наиболее существенно при этом отсутствие классических воспалительных стимулов, приводящих в формированию дендритных клеток типа DC1, а также действие на дендритные клетки IL-10, продуцируемого тучными клетками. В результате формиру-
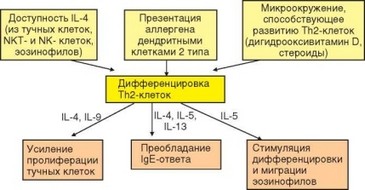
Рис. 4.26. ТИ2-контроль развития аллергии немедленного типа. Условия, благоприятствующие ТИ2-ответу, его продукты (цитокины) и проявления
ются дендритные клетки типа DC2 — продуценты IL-4 (а не IL-12, как при воспалительном иммунном ответе). DC2-клетки мигрируют в региональный лимфатический узел и, презентируя антиген CD4+ Т-лимфоцитам в присутствии IL-4, направляют их развитие по линии Th2-клеток. Недавно выявлены особенности «аллергических» Th2-клеток, главная из которых состоит в секреции наряду с класическими ТИ2-цитокинами провоспалительного цитокина TNFa. Для развития таких клеток необходима секреция цитокина TSLP (Thymic stromal lymphopoietin) эпителиальными клетками слизистых оболочек и кожи. Он индуцирует экспрессию костимулирующей молекулы ОХ40L на дендритных клетках — индукторах Th2-лимфоцитов. Костимуляция через рецептор этой молекулы ОХ40, экспрессируемый Т-клетками, и обеспечивает дифференцировку аллергических ТИ2-клеток.
При последующем поступлении аллергена его презентация дендритными клетками DC2 или В-лимфоцитами Т-клеткам памяти может происходить на месте, обычно в слизистых оболочках. В этом случае в еще большей степени проявляется эффект микроокружения, способствующего диффе- ренцировке Th2-клеток: наличие источников IL-4 (цитокина, который служит основным индуктором Th2-ответа), присутствие стероидов, благоприятствующих дифференцировке Th2-клеток (дигидрооксивитамина D) и т.д. В качестве источника IL-4, необходимого для запуска дифферен- цировки ТИ2-клеток, выступают тучные клетки, NK- и NKT-клетки, а также эозинофилы, если они присутствуют в микроокружении. Перечисленные выше условия наиболее полно реализуются в слизистых оболочках, служащих оптимальным местом развития и реализации аллергических реакций немедленного типа.
Аллергические IgE-антитела
Главная особенность гуморального иммунного ответа на аллергены — более существенный вклад IgE-антител, чем при неаллергическом иммунном ответе. Переключение С-генов иммуноглобулинов на ген Ce, кодирующий
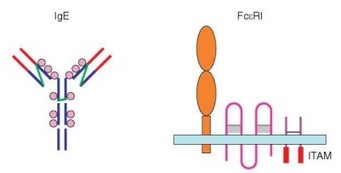
Рис. 4.27. Схема строения молекул IgE и FceRI
изотип IgE, происходит по обычному механизму (см. раздел 3.2.4.3). Выбор гена Ce обусловлен действием IL-4. Переключение осуществляется или одноэтапно (с гена Ср на ген Се) или через стадию переключения на ген Су4, которое также контролируется IL-4. Происходит сближение V- и Ce-генов путем формирования петли ДНК, в которую попадают все гены, локализованные между V- и Ce-генами, с последующим ее вырезанием. Тандем генов V и Ce транскрибируется с образованием мРНК состава VСe с последующей трансляцией Н-цепи IgE.
IgE — 8S мономерный иммуноглобулин (т.е. его молекула содержит 2 тяжелые е- и 2 легкие цепи) с молекулярной массой 190 кДа (рис. 4.27). Характерные особенности структуры е-цепи — наличие четырех С-доменов (вместо трех в у- и a-цепях), обеспечивающее большее пространство для взаимодействия с Fc-рецептором, а также более сильное гликозилирование С-доменов (до 12% углеводов в составе молекулы, 6 сайтов гликозилирова- ния вместо 1—5 в других иммуноглобулинах). Молекула IgE чувствительна к прогреванию при 56 °С, крайним значениям кислотности (рН lt;6 и pH gt;11), восстановлению p-меркаптэтанолом. Вовлечение IgE в развитие аллергии обусловлено наличием в домене Ce3 участка связывания для рецептора FceRI. Некоторый вклад в связывание IgE с FceRI вносит дополнительный сайт, расположенный в домене Се4.
Концентрация IgE в сыворотке крови здорового человека ниже, чем любых других иммуноглобулинов. Она колеблется в пределах 85—350 нг/мл, тогда как при аллергических заболеваниях она может быть на порядки выше. Для определения IgE используют иммуноферментный и радиоиммунный тесты, причем обычно определяют содержание IgE, специфичного к конкретным аллергенам. Содержание IgE выражают в международных единицах — 1 МЕ = 2,42 нг IgE. IgE отсутствует в сыворотке крови новорожденных, но начиная с 3 мес его концентрация постепенно нарастает, достигая уровня взрослых только к 10 годам. Содержание IgE в секретах выше, чем в сыворотке крови (особенно много его в молозиве). Большинство IgE секретируют лимфоидные клетки слизистых оболочек. Сывороточный IgE имеет короткий срок жизни — 2,5 сут.
Выработку IgE контролируют регуляторные факторы. Наиболее специфичен для антител данного изотипа контроль, осуществляемый при участии низкоаффинных Fce-рецепторов (CD23). Этот рецептор экспрессируют В-лимфоциты, а при аллергии — также Т-клетки и моноциты. Под влиянием IL-4 В-клетки и моноциты начинают продуцировать молекулу CD23 в растворимой форме. Взаимодействуя с молекулой CD21 рецепторного комплекса В-лимфоцитов, CD23 запускает сигнал (через тирозинкиназу Lyn, связанную с цитоплазматической частью CD19), способствующий переключению синтеза иммуноглобулинов на IgE, пролиферации IgE+ В-клеток и секреции ими IgE. Выработку IgE стимулируют также ^2-цитокины IL-5 и IL-6.