
Аутотолерантность и ее механизмы
По-видимому, первым поставил вопрос о причинах отсутствия иммунологических реакций на собственные антигены самый проницательный иммунолог рубежа XIX и XX веков П. Эрлих (P. Ehrlich). Не имея рациональных подходов к получению ответа на этот вопрос, он сформулировал аксиоматическую формулу «horror autotoxicus» (боязнь самоотравления). Проблема дискриминации своего и чужого приобрела особую актуальность в 50-е годы прошлого века в связи с доказательством иммунологической природы отторжения трансплантата, открытием феномена иммунологической толерантности и первыми успехами в изучении природы аутоиммунных заболеваний. Необходимость решения этой проблемы породила клонально-селекционную теорию Ф.М. Бернета (F.M. Burnet) — возможно, самую продуктивную теорию в истории иммунологии. В соответствии с одним из основных постулатов этой теории лимфоциты несут на своей поверхности рецепторы для распознавания антигенов, и эти крайне многочисленные рецепторы распределены в популяции лимфоцитов клонально — таким образом, что каждый клон несет рецептор одной специфичности. Другой постулат состоял в том, что характер реакции лимфоцитов на связывание антигена зависит от степени зрелости клетки и организма: незрелые клетки, преобладающие до наступления «иммунологической зрелости» организма, в ответ на контакт с антигеном гибнут, а зрелые активируются, что служит основой иммунного ответа. Именно поэтому до периода иммунологического созревания успевают элиминироваться все клоны лимфоцитов, рецепторы которых распознают собственные антигены организма. Клоны лимфоцитов, распознающие чужеродные (не принадлежащие организму) молекулы, сохраняются и при поступлении этих молекул-антигенов в иммунологически зрелый организм обеспечивают его защиту путем развития иммунного ответа. Ни одно из положений этой теории не опровергнуто, хотя многие уточнены или переформулированы. Следствием этой теории была возможность индукции неотвечаемости (толерантности) к антигенам, вводимым в организм до наступления иммунологической зрелости, что было доказано П. Медаваром (P. Medawar) и его сотрудниками. Им удалось индуцировать иммунологическую толерантность к аллоантигенам (попытка самого Бернета получить толерантность к вирусным антигенам не увенчалась успехом). Таким образом сложилось общепризнанное в настоящее время представление о том, что отсутствие иммунного ответа на собственные антигены является следствием формирования иммунологической толерантности на определенном этапе онтогенеза (в основном — к моменту рождения или первым дням постнатальной жизни).
В настоящее время выделяют активные и пассивные механизмы формирования аутотолерантности. Имеется 4 основных активных механизма формированя аутотолерантности (табл. 4.5, рис. 4.15):
Пассивный механизм аутотолерантности — игнорирование аутоантигенов иммунной системой, обусловленное их низкой концентрацией или изоляцией от иммунной системы.
Таблица 4.5. Механизмы аутотолерантности
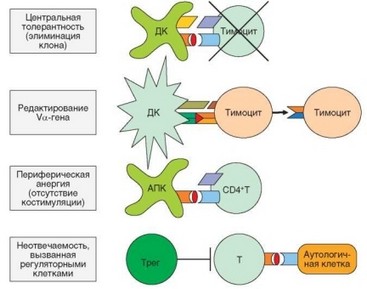
Рис. 4.15. Основные механизмы естественной аутотолерантности. Первый из представленных механизмов реализуется в основном в тимусе, три остальных — в периферическом отделе иммунной системы
Элиминация клонов
В соответствии с современным взглядом на природу аутотолерантности, элиминация аутоспецифических клонов служит ее главным фактором. Элиминация реализуется по механизму отрицательной селекции клонов (см. раздел 3.2.3.4). Особенно подробно изучена отрицательная селекция Т-лимфоцитов. Она осуществляется в тимусе в процессе взаимодействия тимоцитов с клетками стромы. Тимоциты проходят этот этап селекции на стадии CD4+ CD8+, для которой характерен низкий уровень экспрессии антиапоптотических факторов (Bcl-2, Bcl-XL), т.е. в это время клетки высокочувствительны к апоптозу. Селекцию осуществляют 2 типа стромальных клеток — клетки медуллярного эпителия и дендритные клетки тимуса. Ключевое событие селекции — распознавание TCR молекулы MHC, несущей пептидный фрагмент собственных белков-антигенов. Если рецептор обладает высоким сродством к антигенному эпитопу, тимоцит получает сигнал, приводящий к развитию апоптоза клеток. В результате отрицательной селекции из популяции Т-лимфоцитов элиминируются клоны клеток, распознающих антигены, презентируемые в момент селекции в тимусе, т.е. аутоантигены.
Особую проблему составляет степень представленности в тимусе различных антигенов, в особенности внетимусных органоспецифических. Существует особый ген AIRE, ответственный за экспрессию этих анти-
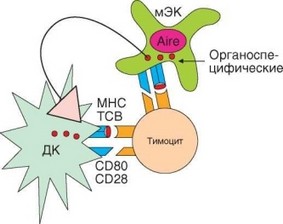
Рис. 4.16. Роль экспрессии гена AIRE в индукции аутотолерантности к органоспецифическим антигенам. Экспрессия гена AIRE вызывает появление в медуллярных эпителиальных клетках тимуса малого количества органоспецифических антигенов нелимфоидных органов. Дифференцирующиеся Т-лимфоциты в процессе отрицательной селекции распознают эти антигены как на самих эпителиальных клетках, так и на дендритных клетках, презентирующих антигены эпителиоцитов. По-видимому, второй путь более эффективен
генов в эпителиальных клетках мозгового слоя тимуса (см. раздел 3.4.1.2). Внетимусные антигены экспрессируются случайным образом в очень ограниченном масштабе, однако достаточном для индукции к ним толерантности (рис. 4.16). Мутации гена AIRE приводят к развитию полиспецифических аутоиммунных процессов.
Наличие отрицательной селекции Т-клеток первоначально было доказано в опытах на мышах путем маркирования групп клонов на принадлежность p-цепей их TCR к различным семействам. Так, клоны Т-клеток, специфичные к некоторым антигенам, несут TCR, содержащие р-цепи определенных семейств. Например, клоны, специфичные к самцовому антигену H-Y, экспрессируют p-цепь семейства Vp6. У самцов, для которых H-Y является аутоантигеном, содержание Vp6+ тимоцитов по мере их созревания снижается, а у самок, для которых Н-У — чужеродный антиген, доля таких тимоцитов, наоборот, возрастает. Особенно яркое доказательство наличия отрицательной селекции — эксперименты с использованием трансгенов (т.е. генов, вводимых в зародышевые клетки). Перенос перестроенных генов TCR известной специфичности приводит к экспрессии соответствующих TCR на всех Т-клетках. При одновременной экспрессии лиганда (антигена и молекул MHC соответствующего аллотипа) в тимусе на стадии CD4+CD8+ тимоцитов элиминируются все Т-клетки. В определенной ситуации, например при низком уровне экспрессии корецептора CD8, некоторые клоны CD8+ Т-клеток избегают элиминации и мигрируют на периферию. Это означает, что существует определенный порог сродства TCR к антигенному лиганду, который должен быть преодолен для осуществления отрицательной селекции.
Отрицательной селекции подвергаются и В-лимфоциты. Ее проходят в костном мозгу и, частично — в периферическом отделе иммунной системы незрелые В-клетки фенотипа IgM+ IgD-, а антигены презентируют стромальные клетки. Аутоспецифические В-клетки погибают вследствие апоптоза. Делеционный характер селекции показан в опытах с двойным переносом генов. Мышам одновременно трансфецировали ген яичного лизоцима и V-гены специфичного к нему BCR. В результате зрелые В-лимфоциты элиминировались при сохранении незрелых В-клеток. Таким образом, отрицательная селекция реализуется на этапе перехода от незрелых В-лимфоцитов к зрелым.
Аутотолерантность, обусловленную элиминацией клонов, распознаю - щих собственные антигены, называют центральной. Она бывает наиболее эффективной при постоянной реализации в течение всей жизни, поскольку популяции Т- и В-лимфоцитов непрерывно обновляются и аутоспецифические клоны могут возникать в течение всей жизни. Как и любой биологический механизм, селекция клонов не достигает 100% эффективности, и единичные аутоспецифические клоны могут пополнять периферический пул лимфоцитов.
Редактирование и анергия
На периферии действует другие механизмы формирования аутотолерантности и предотвращения аутоагрессии. Распознавание аутоантигена в периферическом отделе иммунной системы служит сигналом для редактирования гена a-цепи TCR. Вновь экспрессируются гены RAG и осуществляется повторная перестройка гена TRAV, обычно приводящая к изменению специфичности TCR и утрате ею аутоспецифичности.
Если аутоспецифичность сохраняется при эмиграции Т-клетки их тимуса, индуцируется ее анергия, т.е. неотвечаемость. Условием ее формирования является отсутствие костимуляции при распознавании Т-клеткой аутоантигена (рис. 4.17). Это соответствует ситуации, когда антиген представляется Т-лимфоциту непрофессиональной АПК, лишенной костимули- рующих молекул CD80 и CD86, способных осуществить дополнительную сигнализацию через молекулу CD28 (см. раздел 3.5.1.4). Если Т-лимфоцит распознает антиген на АПК в отсутствие костимуляции, полноценный активирующий сигнал не формируется, и клетка подвергается анергии. Это явление специфично — анергии подвергаются конкретные клоны Т-лимфоцитов.
Механизмы анергии раскрыты не полностью. Установлено, что формирование этого состояния сопряжено с ослабением активности рецепторной тирозинкиназы Lck, отсутствием или ослаблением фосфорилирования киназы ZAP-70 и адапторного белка LAT. Среди низкомолекулярных ГТФаз в этот поцесс в наибольшей степени вовлечен фактор р21ras. В связи с этим из основных сигнальных путей, запускаемых в клетке при актвиации через TCR/CD28, сильнее всего страдает MAP-каскад и особенно та его ветвь, которая зависит от Ras. Она приводит к формированию сигнальных факторов ERK и c-Fos. Однако активность JNK- и p3 8-ветвей MAP-каскада
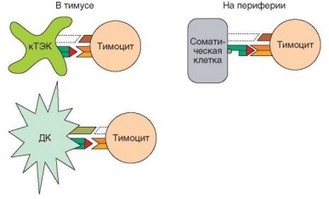
Рис. 4.17. Механизмы индукции анергии Т-лимфоцитов в тимусе и периферическом отделе иммунной системы. Главный механизм индукции анергии Т-клеток — отсутствие одного из компонентов костимулирующей системы в момент распознавания антигена. Может иметь значение отсутствие костимулирующих молекул (В7) на кортикальных эпителиальных клетках тимуса или соматических клетках вне тимуса или отсутствие молекул CD28 на созревающих тимоцитах
также ослабляется. В результате из трех основных транскрипционных факторов, активируемых при презентации антигена, в наибольшей степени нарушается образование фактора АР-1 (димера c-Fos/c-Jun). Среди генов, активируемых при передаче сигнала через TCR/CD28, сильнее всего страдает экспрессия гена IL-2, наиболее важного для активации Т-клеток. Процессы анергии и активации различаются не только перечисленными особенностями передачи сигнала. Так, показана роль фактора Cbl-b в качестве ингибитора активации, а также вклад ряда других факторов в индукцию анергии. Обычным исходом анергии является ускоренная гибель клетки по механизму апоптоза.
Реальность анергии Т-лимфоцитов иллюстрирует следующий эксперимент. Мышам генотипа Н-2а трансфецировали гены Н-2ь и одновременно — гены TCR, специфичного к Н-2ь, таким образом, чтобы ген H-2b экспрессировался в клетках поджелудочной железы, а анти-H-2b TCR — в Т-лим- фоцитах. Тимоциты таких мышей реагировали на стимуляцию Н-2ь образованием специфических Т-киллеров, тогда как периферические Т-клетки не реагировали на H-2b. Следовательно, у таких мышей клон анти-Н-2ь Т-клеток не элиминировался в тимусе, но его реакция на эту молекулу MHC на периферии была подавлена. С помощью дополнительных экспериментов было показано по крайней мере временное присутствие на периферии ареак- тивных Т-клеток, специфичных к Н-2Ь. Таким образом можно продемонстрировать существование периферической толерантности, обусловленной не элиминацией, а анергией Т-клона.
Индукция анергии В-лимфоцитов более проблематична. Однако в соответствии с закономерностями иммунного ответа для предотвращения акти
вации клонов В-лимфоцитов бывает достаточно элиминации или индукции анергии клонов Т-клеток той же специфичности.
Роль регуляторных Т-клеток
Однако индукции периферической толерантности по механизму анергии оказывается недостаточно для надежного предотвращения аутоагрессии. Существует еще один — третий механизм аутотолерантности, который опосредован регуляторными Т-клетками. В связи с активным характером такой толерантности и возможностью ее переноса сингенным реципиентам ее называют доминантной (толерантность, обусловленная элиминацией или анергией клонов, не только не может быть перенесена, но и устраняется при переносе интактных лимфоцитов в связи с присутствием в них реактивных клонов). Еще в 70-е годы было постулировано наличие в иммунной системе особой разновидности супрессорных Т-лимфоцитов, «запрещающих» реакцию на собственные антигены. Такие клетки назвали вето-клетками.
Реальность их существования была показана только в 1990-е годы, когда были описаны естественные регуляторные Т-клетки. Эти клетки, имеющие мембранный фенотип CD4+ CD25hi CTLA-4+ и экспрессирующие внутриклеточный дифференцировочный фактор FOXP3, охарактеризованы выше (см. раздел 3.3.2.5). Особенность развития этих клеток состоит в том, что они в значительной степени избегают отрицательной селекции. Порог сродства к аутоантигену, обеспечивающий запуск летального сигнала, для этих клеток выше, чем для всех остальных Т-лимфоцитов. В связи с этим некоторые CD4+ Т-клетки, специфичные к аутоантигенам, не подвергаются апоптозу и дифференцируются в регуляторные Т-лимфоциты. Они эмигрируют на периферию, их выявляют в лимфатических узлах и других лимфоидных тканях, эти клетки препятствуют активации аутоспецифических эффекторных Т-лимфоцитов, если таковые избежали элиминации и анергии на других этапах индукции толерантности. Таким образом, аутоспецифичность регуляторных Т-клеток не только не перерастает в аутоагрессию, но, наоборот, служит условием ее предотвращения.
Игнорирование
Суть игнорирования состоит в том, что антиген не может вызвать реакцию иммунной системы, если его концентрация в организме ниже пороговой (порог различен для разных молекул). Наличие такого порога важно и для формирования аутотолерантности, и для индукции иммунного ответа. Обычно аутотолерантность не формируется по отношению к молекулам, присутствующим в организме в очень низких концентрациях, однако при этом иммунный ответ также не развивается. В некоторых случаях концентрация таких веществ может возрастать. Это может происходить при опухолевом росте, приводящем к увеличению экспрессии опухолеассоциированного антигена, до того вырабатывавшегося немногочисленными клетками, или при иммунном ответе, когда значительно нарастает концентрация антител, несущих определенный идиотип. В этих ситуациях может развиться иммунный ответ на подобные антигены, к которым аутотолерантность не сформировалась. Вариант игнорирования — отсутствие реакции иммунной системы на антигены, изолированные от нее тканевыми барьерами, как это бывает в иммунологически привилегированных органах (см. раздел 4.3.2.3).
В этом случае нарушение барьера может привести к развитию аутоиммунного процесса.
Таким образом, аутотолерантность формируется с помощью трех активных механизмов — элиминации клонов в ходе отрицательной селекции (центральная толерантность), анергии клонов в периферическом отделе иммунной системы (периферическая толерантность) и контроля со стороны аутоспецифичных регуляторных Т-лимфоцитов. Важен также феномен игнорирования антигенов, присутствующих в слишком низких концентрациях или недоступных для распознавания иммунной системой. Относительный вклад этих механизмов в предотвращение аутоагрессии оценить трудно. По-видимому, все они обязательны для поддержания аутотолерантности. Отключение процесса отрицательной селекции и предотвращение развития регуляторных клеток неизбежно приводят к развитию фатальных полиспецифических аутоиммунных процессов. Нарушение аутотолерантности служит основой аутоиммунных заболеваний — одного из основных проявлений иммунопатологии.
По-видимому, первым поставил вопрос о причинах отсутствия иммунологических реакций на собственные антигены самый проницательный иммунолог рубежа XIX и XX веков П. Эрлих (P. Ehrlich). Не имея рациональных подходов к получению ответа на этот вопрос, он сформулировал аксиоматическую формулу «horror autotoxicus» (боязнь самоотравления). Проблема дискриминации своего и чужого приобрела особую актуальность в 50-е годы прошлого века в связи с доказательством иммунологической природы отторжения трансплантата, открытием феномена иммунологической толерантности и первыми успехами в изучении природы аутоиммунных заболеваний. Необходимость решения этой проблемы породила клонально-селекционную теорию Ф.М. Бернета (F.M. Burnet) — возможно, самую продуктивную теорию в истории иммунологии. В соответствии с одним из основных постулатов этой теории лимфоциты несут на своей поверхности рецепторы для распознавания антигенов, и эти крайне многочисленные рецепторы распределены в популяции лимфоцитов клонально — таким образом, что каждый клон несет рецептор одной специфичности. Другой постулат состоял в том, что характер реакции лимфоцитов на связывание антигена зависит от степени зрелости клетки и организма: незрелые клетки, преобладающие до наступления «иммунологической зрелости» организма, в ответ на контакт с антигеном гибнут, а зрелые активируются, что служит основой иммунного ответа. Именно поэтому до периода иммунологического созревания успевают элиминироваться все клоны лимфоцитов, рецепторы которых распознают собственные антигены организма. Клоны лимфоцитов, распознающие чужеродные (не принадлежащие организму) молекулы, сохраняются и при поступлении этих молекул-антигенов в иммунологически зрелый организм обеспечивают его защиту путем развития иммунного ответа. Ни одно из положений этой теории не опровергнуто, хотя многие уточнены или переформулированы. Следствием этой теории была возможность индукции неотвечаемости (толерантности) к антигенам, вводимым в организм до наступления иммунологической зрелости, что было доказано П. Медаваром (P. Medawar) и его сотрудниками. Им удалось индуцировать иммунологическую толерантность к аллоантигенам (попытка самого Бернета получить толерантность к вирусным антигенам не увенчалась успехом). Таким образом сложилось общепризнанное в настоящее время представление о том, что отсутствие иммунного ответа на собственные антигены является следствием формирования иммунологической толерантности на определенном этапе онтогенеза (в основном — к моменту рождения или первым дням постнатальной жизни).
В настоящее время выделяют активные и пассивные механизмы формирования аутотолерантности. Имеется 4 основных активных механизма формированя аутотолерантности (табл. 4.5, рис. 4.15):
- элиминация аутоспецифических клонов (центральный механизм аутотолерантности);
- редактиврование генов аутоспецифических рецепторов;
- индукция анергии аутоспецифических клонов (периферический механизм аутотолерантности);
- подавление аутоспецифического ответа регуляторными клетками (доминантный механизм аутотолерантности).
Пассивный механизм аутотолерантности — игнорирование аутоантигенов иммунной системой, обусловленное их низкой концентрацией или изоляцией от иммунной системы.
Таблица 4.5. Механизмы аутотолерантности
|
Механизм |
Путь реализации |
Место реализации |
|
Делеция клонов |
В процессе отрицательной селекции медуллярные эпителиальные и дендритные клетки тимуса индуцируют апоптоз Т-клеток, несущих TCR, обладающий высоким сродством к аутоантигенам. Полнота элиминации клонов дополнительно обеспечивается экспрессией в тимусе органоспецифических антигенов нелимфоидных органов (под контролем гена AIRE). Аутоспецифические В-клетки элиминируются в результате аналогичного взаимодействия со стромальными клетками |
Для Т-клеток — тимус (кортикомедуллярная зона и мозговой слой). Для В-кле- ток — костный мозг |
|
Редактирование рецепторных генов |
При распознавании незрелыми клетками аутоантигенов запускается повторная перестройка Va-гена TCR и VK/X-генов Ig |
Вторичные лимфоидные органы |
|
Анергия |
При распознавании аутоантигена, не поддерживаемого кости- муляцией, происходит стабильная утрата способности клетки к активации |
Нелимфоидные органы |
|
Контроль со стороны регуляторных T-клеток |
При одновременной презентации дендритной клеткой аутоантигена регуляторной и эффек- торной Т-клеткам, регуляторные Т-лимфоциты подавляют ответ эффек- торных Т-клеток по контактному механизму. Клетки типов Th3 и Tr1 подавляют ответ с помощью гуморальных факторов — TGF р и IL-10 |
Вторичные лимфоидные органы |
Рис. 4.15. Основные механизмы естественной аутотолерантности. Первый из представленных механизмов реализуется в основном в тимусе, три остальных — в периферическом отделе иммунной системы
Элиминация клонов
В соответствии с современным взглядом на природу аутотолерантности, элиминация аутоспецифических клонов служит ее главным фактором. Элиминация реализуется по механизму отрицательной селекции клонов (см. раздел 3.2.3.4). Особенно подробно изучена отрицательная селекция Т-лимфоцитов. Она осуществляется в тимусе в процессе взаимодействия тимоцитов с клетками стромы. Тимоциты проходят этот этап селекции на стадии CD4+ CD8+, для которой характерен низкий уровень экспрессии антиапоптотических факторов (Bcl-2, Bcl-XL), т.е. в это время клетки высокочувствительны к апоптозу. Селекцию осуществляют 2 типа стромальных клеток — клетки медуллярного эпителия и дендритные клетки тимуса. Ключевое событие селекции — распознавание TCR молекулы MHC, несущей пептидный фрагмент собственных белков-антигенов. Если рецептор обладает высоким сродством к антигенному эпитопу, тимоцит получает сигнал, приводящий к развитию апоптоза клеток. В результате отрицательной селекции из популяции Т-лимфоцитов элиминируются клоны клеток, распознающих антигены, презентируемые в момент селекции в тимусе, т.е. аутоантигены.
Особую проблему составляет степень представленности в тимусе различных антигенов, в особенности внетимусных органоспецифических. Существует особый ген AIRE, ответственный за экспрессию этих анти-
Рис. 4.16. Роль экспрессии гена AIRE в индукции аутотолерантности к органоспецифическим антигенам. Экспрессия гена AIRE вызывает появление в медуллярных эпителиальных клетках тимуса малого количества органоспецифических антигенов нелимфоидных органов. Дифференцирующиеся Т-лимфоциты в процессе отрицательной селекции распознают эти антигены как на самих эпителиальных клетках, так и на дендритных клетках, презентирующих антигены эпителиоцитов. По-видимому, второй путь более эффективен
генов в эпителиальных клетках мозгового слоя тимуса (см. раздел 3.4.1.2). Внетимусные антигены экспрессируются случайным образом в очень ограниченном масштабе, однако достаточном для индукции к ним толерантности (рис. 4.16). Мутации гена AIRE приводят к развитию полиспецифических аутоиммунных процессов.
Наличие отрицательной селекции Т-клеток первоначально было доказано в опытах на мышах путем маркирования групп клонов на принадлежность p-цепей их TCR к различным семействам. Так, клоны Т-клеток, специфичные к некоторым антигенам, несут TCR, содержащие р-цепи определенных семейств. Например, клоны, специфичные к самцовому антигену H-Y, экспрессируют p-цепь семейства Vp6. У самцов, для которых H-Y является аутоантигеном, содержание Vp6+ тимоцитов по мере их созревания снижается, а у самок, для которых Н-У — чужеродный антиген, доля таких тимоцитов, наоборот, возрастает. Особенно яркое доказательство наличия отрицательной селекции — эксперименты с использованием трансгенов (т.е. генов, вводимых в зародышевые клетки). Перенос перестроенных генов TCR известной специфичности приводит к экспрессии соответствующих TCR на всех Т-клетках. При одновременной экспрессии лиганда (антигена и молекул MHC соответствующего аллотипа) в тимусе на стадии CD4+CD8+ тимоцитов элиминируются все Т-клетки. В определенной ситуации, например при низком уровне экспрессии корецептора CD8, некоторые клоны CD8+ Т-клеток избегают элиминации и мигрируют на периферию. Это означает, что существует определенный порог сродства TCR к антигенному лиганду, который должен быть преодолен для осуществления отрицательной селекции.
Отрицательной селекции подвергаются и В-лимфоциты. Ее проходят в костном мозгу и, частично — в периферическом отделе иммунной системы незрелые В-клетки фенотипа IgM+ IgD-, а антигены презентируют стромальные клетки. Аутоспецифические В-клетки погибают вследствие апоптоза. Делеционный характер селекции показан в опытах с двойным переносом генов. Мышам одновременно трансфецировали ген яичного лизоцима и V-гены специфичного к нему BCR. В результате зрелые В-лимфоциты элиминировались при сохранении незрелых В-клеток. Таким образом, отрицательная селекция реализуется на этапе перехода от незрелых В-лимфоцитов к зрелым.
Аутотолерантность, обусловленную элиминацией клонов, распознаю - щих собственные антигены, называют центральной. Она бывает наиболее эффективной при постоянной реализации в течение всей жизни, поскольку популяции Т- и В-лимфоцитов непрерывно обновляются и аутоспецифические клоны могут возникать в течение всей жизни. Как и любой биологический механизм, селекция клонов не достигает 100% эффективности, и единичные аутоспецифические клоны могут пополнять периферический пул лимфоцитов.
Редактирование и анергия
На периферии действует другие механизмы формирования аутотолерантности и предотвращения аутоагрессии. Распознавание аутоантигена в периферическом отделе иммунной системы служит сигналом для редактирования гена a-цепи TCR. Вновь экспрессируются гены RAG и осуществляется повторная перестройка гена TRAV, обычно приводящая к изменению специфичности TCR и утрате ею аутоспецифичности.
Если аутоспецифичность сохраняется при эмиграции Т-клетки их тимуса, индуцируется ее анергия, т.е. неотвечаемость. Условием ее формирования является отсутствие костимуляции при распознавании Т-клеткой аутоантигена (рис. 4.17). Это соответствует ситуации, когда антиген представляется Т-лимфоциту непрофессиональной АПК, лишенной костимули- рующих молекул CD80 и CD86, способных осуществить дополнительную сигнализацию через молекулу CD28 (см. раздел 3.5.1.4). Если Т-лимфоцит распознает антиген на АПК в отсутствие костимуляции, полноценный активирующий сигнал не формируется, и клетка подвергается анергии. Это явление специфично — анергии подвергаются конкретные клоны Т-лимфоцитов.
Механизмы анергии раскрыты не полностью. Установлено, что формирование этого состояния сопряжено с ослабением активности рецепторной тирозинкиназы Lck, отсутствием или ослаблением фосфорилирования киназы ZAP-70 и адапторного белка LAT. Среди низкомолекулярных ГТФаз в этот поцесс в наибольшей степени вовлечен фактор р21ras. В связи с этим из основных сигнальных путей, запускаемых в клетке при актвиации через TCR/CD28, сильнее всего страдает MAP-каскад и особенно та его ветвь, которая зависит от Ras. Она приводит к формированию сигнальных факторов ERK и c-Fos. Однако активность JNK- и p3 8-ветвей MAP-каскада
Рис. 4.17. Механизмы индукции анергии Т-лимфоцитов в тимусе и периферическом отделе иммунной системы. Главный механизм индукции анергии Т-клеток — отсутствие одного из компонентов костимулирующей системы в момент распознавания антигена. Может иметь значение отсутствие костимулирующих молекул (В7) на кортикальных эпителиальных клетках тимуса или соматических клетках вне тимуса или отсутствие молекул CD28 на созревающих тимоцитах
также ослабляется. В результате из трех основных транскрипционных факторов, активируемых при презентации антигена, в наибольшей степени нарушается образование фактора АР-1 (димера c-Fos/c-Jun). Среди генов, активируемых при передаче сигнала через TCR/CD28, сильнее всего страдает экспрессия гена IL-2, наиболее важного для активации Т-клеток. Процессы анергии и активации различаются не только перечисленными особенностями передачи сигнала. Так, показана роль фактора Cbl-b в качестве ингибитора активации, а также вклад ряда других факторов в индукцию анергии. Обычным исходом анергии является ускоренная гибель клетки по механизму апоптоза.
Реальность анергии Т-лимфоцитов иллюстрирует следующий эксперимент. Мышам генотипа Н-2а трансфецировали гены Н-2ь и одновременно — гены TCR, специфичного к Н-2ь, таким образом, чтобы ген H-2b экспрессировался в клетках поджелудочной железы, а анти-H-2b TCR — в Т-лим- фоцитах. Тимоциты таких мышей реагировали на стимуляцию Н-2ь образованием специфических Т-киллеров, тогда как периферические Т-клетки не реагировали на H-2b. Следовательно, у таких мышей клон анти-Н-2ь Т-клеток не элиминировался в тимусе, но его реакция на эту молекулу MHC на периферии была подавлена. С помощью дополнительных экспериментов было показано по крайней мере временное присутствие на периферии ареак- тивных Т-клеток, специфичных к Н-2Ь. Таким образом можно продемонстрировать существование периферической толерантности, обусловленной не элиминацией, а анергией Т-клона.
Индукция анергии В-лимфоцитов более проблематична. Однако в соответствии с закономерностями иммунного ответа для предотвращения акти
вации клонов В-лимфоцитов бывает достаточно элиминации или индукции анергии клонов Т-клеток той же специфичности.
Роль регуляторных Т-клеток
Однако индукции периферической толерантности по механизму анергии оказывается недостаточно для надежного предотвращения аутоагрессии. Существует еще один — третий механизм аутотолерантности, который опосредован регуляторными Т-клетками. В связи с активным характером такой толерантности и возможностью ее переноса сингенным реципиентам ее называют доминантной (толерантность, обусловленная элиминацией или анергией клонов, не только не может быть перенесена, но и устраняется при переносе интактных лимфоцитов в связи с присутствием в них реактивных клонов). Еще в 70-е годы было постулировано наличие в иммунной системе особой разновидности супрессорных Т-лимфоцитов, «запрещающих» реакцию на собственные антигены. Такие клетки назвали вето-клетками.
Реальность их существования была показана только в 1990-е годы, когда были описаны естественные регуляторные Т-клетки. Эти клетки, имеющие мембранный фенотип CD4+ CD25hi CTLA-4+ и экспрессирующие внутриклеточный дифференцировочный фактор FOXP3, охарактеризованы выше (см. раздел 3.3.2.5). Особенность развития этих клеток состоит в том, что они в значительной степени избегают отрицательной селекции. Порог сродства к аутоантигену, обеспечивающий запуск летального сигнала, для этих клеток выше, чем для всех остальных Т-лимфоцитов. В связи с этим некоторые CD4+ Т-клетки, специфичные к аутоантигенам, не подвергаются апоптозу и дифференцируются в регуляторные Т-лимфоциты. Они эмигрируют на периферию, их выявляют в лимфатических узлах и других лимфоидных тканях, эти клетки препятствуют активации аутоспецифических эффекторных Т-лимфоцитов, если таковые избежали элиминации и анергии на других этапах индукции толерантности. Таким образом, аутоспецифичность регуляторных Т-клеток не только не перерастает в аутоагрессию, но, наоборот, служит условием ее предотвращения.
Игнорирование
Суть игнорирования состоит в том, что антиген не может вызвать реакцию иммунной системы, если его концентрация в организме ниже пороговой (порог различен для разных молекул). Наличие такого порога важно и для формирования аутотолерантности, и для индукции иммунного ответа. Обычно аутотолерантность не формируется по отношению к молекулам, присутствующим в организме в очень низких концентрациях, однако при этом иммунный ответ также не развивается. В некоторых случаях концентрация таких веществ может возрастать. Это может происходить при опухолевом росте, приводящем к увеличению экспрессии опухолеассоциированного антигена, до того вырабатывавшегося немногочисленными клетками, или при иммунном ответе, когда значительно нарастает концентрация антител, несущих определенный идиотип. В этих ситуациях может развиться иммунный ответ на подобные антигены, к которым аутотолерантность не сформировалась. Вариант игнорирования — отсутствие реакции иммунной системы на антигены, изолированные от нее тканевыми барьерами, как это бывает в иммунологически привилегированных органах (см. раздел 4.3.2.3).
В этом случае нарушение барьера может привести к развитию аутоиммунного процесса.
Таким образом, аутотолерантность формируется с помощью трех активных механизмов — элиминации клонов в ходе отрицательной селекции (центральная толерантность), анергии клонов в периферическом отделе иммунной системы (периферическая толерантность) и контроля со стороны аутоспецифичных регуляторных Т-лимфоцитов. Важен также феномен игнорирования антигенов, присутствующих в слишком низких концентрациях или недоступных для распознавания иммунной системой. Относительный вклад этих механизмов в предотвращение аутоагрессии оценить трудно. По-видимому, все они обязательны для поддержания аутотолерантности. Отключение процесса отрицательной селекции и предотвращение развития регуляторных клеток неизбежно приводят к развитию фатальных полиспецифических аутоиммунных процессов. Нарушение аутотолерантности служит основой аутоиммунных заболеваний — одного из основных проявлений иммунопатологии.