
В основе всех событий, происходящих в сенсибилизированном организме при повторном поступлении аллергена, лежит реакция тучных клеток на связывание аллергена с антителами, фиксированными на их поверхности (рис. 4.28).
Тучные клетки и аллергия
Подробная характеристика тучных клеток дана ранее (см. раздел 2.1.4). Их очень много в барьерных тканях, особенно в слизистых оболочках. В развитии тучных клеток, помимо основного фактора SCF, участвуют цитокины, секретируемые ^2-лимфоцитами и самими тучными клетками — IL-4, IL-3, IL-9, IL-10. Тучные клетки участвуют в запуске воспалительной реакции и выступают в качестве эффекторных клеток в защите от макропаразитов. При патологии они оказываются основными эффекторными клетками аллергических реакций немедленного типа. Участие тучных клеток в развитии таких реакций многообразно. В частности, они вносят важный вклад в формирование фона, благоприятствующего развитию Th2-зависимого иммунного ответа и привлечение дополнительных эффекторов аллергии (комплекс секретируемых ими цитокинов близок таковому ^2-клеток и включает IL-4, Il-5, IL-10 и другие ^2-цитокины). Однако наиболее значимо для реализации основного события аллергической реакции — выброса гистамина и других активных молекул — наличие на поверхности этих клеток рецепторов FceRI и способность тучных клеток отвечать дегрануляцией на перекрестное связывание этих рецепторов (см. рис. 4.28).
Рецепторы FceRI охарактеризованы выше (см. раздел 2.3.4.2.). Они содержат цепи трех типов — a-цепь, связывающую IgE-антитела, р-цепь (4-кратно пронизывающую мембрану) и сигнальную у-цепь, структурно родственную Z-цепи TCR и подобно ей содержащую 3 активационные последовательности ITAM (см. рис. 4.27). Связывание с рецептором свободных молекул IgE-антител, возможное благодаря высокому сродству их Fc-части к рецептору (Kd = 10-10 М), не вызывает реакции со стороны тучных клеток. Реакция возникает при перекрестном сшивании рецепторов благодаря бивалентности аллергена. Только в этом случае происходят конформацион- ные изменения a-цепи, улавливаемые связанной с ней тирозинкиназой Lyn, которая при этом фосфорилируется и активируется. Под влиянием этой
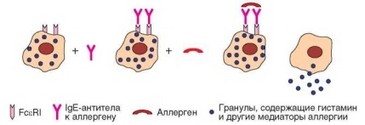
Рис. 4.28. Центральная роль тучных клеток в механизмах немедленной реакции гиперчувствительности. На рисунке представлена роль в развитии аллергии тучных клеток, IgE-антител и аллергена
киназы сигнал передается на в- и у-цепи. В последней фосфорилируются все 3 ITAM. Это необходимо для передачи сигнала тирозинкиназе Syk, что аналогично передаче активационного сигнала в лимфоцитах. Как известно, ключевое событие передачи сигнала в Т-клетке — взваимодействие Z-цепи рецептора (имеющей 3 участка ITAM) и киназы ZAP-70, относящейся к семейству Syk; в В-клетке происходят сходные события, в которых участвует киназа Syk.
На следующем этапе активации (как и при активации лимфоцитов) участвует ряд ферментов, ГТФаз и малых ГТФ-связывающих белков, а затем происходит разделение единого сигнального пути на 3 ветви, приводящие к разным результатам, которые обеспечивают реализацию основных последствий действия аллергена.
Один из ферментов, активируемых тирозинкиназой Syk, — у-изоформа фосфолипазы С (PLCy). Она катализирует расщепление фосфатдилинози- тол дифосфата на диацилглицерол и инозитол-3-фосфат (см. раздел 3.5.2.1). При этом в тучных клетках происходит повышение внутриклеточной концентрации ионов Са2+ и активация протеинкиназы С. Этот фермент фос- форилирует L-цепи миозина и участвует в других процессах, обусловливающих реакцию сократительных элементов цитоскелета и приводящих к выбросу гранул — дегрануляции.
Другой фермент, активирующийся под влиянием киназы Syk (при участии белка Ras и ионов Са2+, накапливающих при реализации параллельных сигнальных путей) — фосфолипаза A2 (PLA2). Действуя на фосфорилхолин, она катализирует синтез арахидоновой кислоты, из которой образуются эйкозаноиды (см. раздел 2.5.4), секретируемые вскоре после дегрануляции. Наконец, запуск Ras-зависимого MAP-каскада приводит к формированию транскрипционного фактора АР-1, включающего гены цитокинов (рис. 4.29).
Дегрануляция тучных клеток, действие предобразованных факторов и эйкозаноидов
Дегрануляция — типовая реакция тучных клеток, их ответ на стимуляцию. Изменения морфологии клеток, связанные с дегрануляцией, представлены на рис. 4.30. Причиной дегрануляции может быть не только связывание аллергена с комплексом IgE/FceRI, но и другие воздействия,
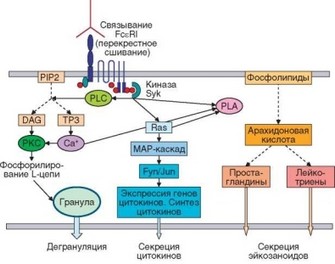
Рис. 4.29. Сигнальные пути и процессы, ответственные за осуществление реакций гиперчувствительности немедленного типа. Через рецептор FceRI в клетку поступает сигнал, реализуемый в виде нескольких сигнальных каскадов. Результаты проявляются в разное время: сначала происходит дегрануляция тучных клеток, затем — синтез эйкозаноидов и еще позже — секреция цитокинов. Эти факторы определяют последовательную смену процессов, формирующих аллергическую реакцию
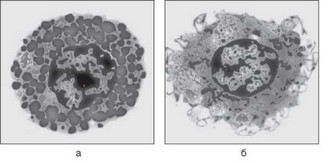
Рис. 4.30. Тучные клетки: а — покоящаяся; б — подвергшаяся дегрануляции. Микрофотографии
приводящие к росту внутриклеточной концентарции Са2+. К ним относят связывание анафилатоксинов (например, С5а) с рецепторами тучных клеток, влияние кальциевых ионофоров и фармакологических агентов (например, холинергических), повышающих уровень Са2+ в цитозоле.
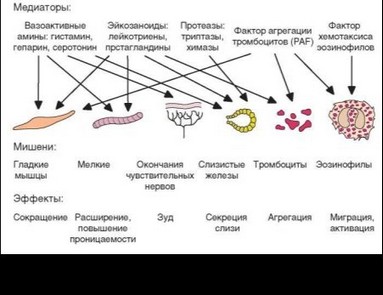
Быстрота развития аллергии немедленного типа обусловлена наличием в гранулах предобразованных факторов, которые проявляют свое действие немедленно после выхода в межклеточное пространство (табл. 4.10). Гранулы тучных клеток содержат вазоактивные амины, включая главный из них — гистамин, а также пептидогликаны — хондроитинсульфаты А и С и гепарин (их соотношение варьирует между субпопуляциями тучных клеток), ферменты (протеазы, дегидрогеназа, пероксидаза, РНКаза, гисти- динкарбоксилаза и кислые гликозамингликаны).
Таблица 4.10. Характеристика факторов, содержащиеся в гранулах тучных клеток (первичных медиаторов аллергии)
|
Фактор |
Химическая природа |
Эффекты |
|
Гистамин |
Амин, метаболит гистидина |
Расширение и повышение проницаемости сосудов, спазм гладкой мускулатуры, усиление секреции слизи, раздражение нервных окончаний (зуд) |
|
Гепарин |
Пептидогли- кан |
Расширение и повышение проницаемости сосудов, спазм гладкой мускулатуры |
|
Серотонин |
Амин, метаболит триптофана |
Расширение и повышение проницаемости сосудов, спазм гладкой мускулатуры |
|
Химаза, триптаза |
Белки |
Протеолиз, усиление секреции слизи, ремоделирование эпителия |
|
Хемотаксический фактор эозинофилов |
Белок |
Хемотаксис эозинофилов |
|
Хемотаксический фактор нейтрофилов |
Белок |
Хемотаксис нейтрофилов |
Главный эффекторный фактор ранней фазы гиперчувствительности немедленного типа — гистамин (см. раздел 2.5.3). Он действует на клетки, экспрессирующие рецепторы для этого амина. Н1-рецепторы представлены на гладких мышцах и эндотелии сосудов, что обеспечивает два главных эффекта гистамина — спазм гладкой мускулатуры бронхов и расширение сосудов с повышением проницаемости капилляров. С действием гистамина связано также усиление секреции слизи и ощущение зуда, характерного для аллергических реакций. Через Н2-рецепторы нейтрофилов и лимфоцитов реализуются регуляторные эффекты гистамина.
Гепарин и другие пептидогликаны также обусловливают основные проявления ранней фазы аллергической реакции — расширение сосудов, повышение их проницаемости и спазм гладкой мускулаторы, но выраженность их эффектов слабее, чем у гистамина. Протеазы (химаза и триптаза) вызывают иные эффекты, характерные для более поздней фазы аллергической реакции, — они обусловливают локальный протеолиз, усиление секреции слизи, ремоделирование эпителия.
Вторая волна выделения активных молекул активированными тучными клетками (табл. 4.11) связана с секрецией эйкозаноидов (см. рис. 4.29).
Эйкозаноиды — липидные метаболиты, производные арахидоновой кислоты, служащие медиаторами аллергических реакций. Их подробная характеристика представлена выше (см. раздел 2.5.4). К активным участникам аллергичеких реакций относят лейкотриены LTC4, LTD4, LTE4 и проста- гландин D2 (в меньшей степени E2, Fa2 и I2), а также тромбоксан TxA2;. Для большинства перечисленных эйкозаноидов клетками-мишенями служат эндотелиальные клетки сосудов, гладкие мышцы, особенно бронхиальные, а также клетки слизистых желез, тромбоциты и нейтрофилы. Действуя через рецепторы, эти факторы, прежде всего лейкоториены, расширяют сосуды (тромбоксан суживает их) и повышают их проницаемость, вызывают спазм гладких мышц (основной эффект — бронхоспазм), стимулируют выделение слизи, вызывают хемотаксис нейтрофилов. Среди эффектов лейкотриенов доминирует их действие на гладкие мышцы бронхов: они обусловливают реализацию медленной фазы бронхоспазма. Простагландины (особенно простагландин E2) оказывают иммунорегуляторное (преимущественно су- прессирующее) действие. Тромбоксан А2 вызывает агрегацию тромбоцитов с освобождением из них ферментов и других активных факторов, способствующих пролиферации лимфоцитов.
Таблица 4.11. Гуморальные факторы, высвобождаемые эффекторными клетками аллергии
|
Группа факторов |
Тучные клетки |
Эозинофилы |
|
Предобразованные факторы |
Гистамин, гепарин, хондроитинсульфат, серотонин, протеазы |
Главный основный белок (МВР), катионный белок эозинофилов (ЕСР), пероксидаза эозинофилов (EPO), нейротоксин эозинофилов (EDT) |
|
Быстро синтезируемые факторы |
Лейкотриены (LTC4, LTD4), простагландины (D2), тромбоксаны, фактор, активирующий тромбоциты (PAF) |
Лейкотриены (LTC4, LTD4), про- стагландины (E), фактор, активирующий тромбоциты (PAF) |
|
Медленно синтезируемые факторы |
IL-5, GM-CSF, TNFa, IL-8 |
IL-3, IL-5, GM-CSF, TGFp |
Поздняя фаза аллергии немедленного типа
Отложенная фаза немедленной гиперчувствительности развивается позже 4—6 ч после действия аллергена (рис. 4.31). Как сказано выше, она обусловлена привлечением из циркуляции крови эозинофилов, базофилов и нейтрофилов цитокинами, синтезируемыми тучными клетками и Th2-лимфоцитами, — IL-4, IL-5, IL-9, IL-13, а также хемокинами. Названные цитокины и гуморальные продукты мигрирующих клеток, в первую очередь эозинофилов, определяют характерные черты этой фазы развития аллергической реакции.
Активация мигрирующих клеток провоспалительными цитокинами, хемокинами, анафилатоксинами и другими факторами, присутствующими в очаге аллергического поражения, приводит к их дегрануляции и секреции ими цитокинов (рис. 4.32). Основные факторы, выделяемые эозинофила-
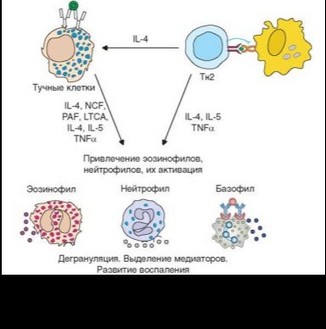
ми — главные компоненты их специфических гранул: главный щелочной белок (MBP), эозинофильный катионный белок (ECP), эозинофильная пероксидаза (ЕРО) и нейротоксин эозинофилов (EDN) (см. раздел 2.1.3). Эти факторы играют важную роль в защите от гельминтов, однако их действие при аллергии изучено неполно. Установлено, что МВР стимулирует тучные клетки и базофилы и вносит вклад в формирование гиперреактивности бронхов. Все вышеперечисленные белки эозинофилов в силу своей цитотоксичности участвуют в повреждении тканей в очаге поражения. Эозинофилы продуцируют также эйкозаноиды, прежде всего LTC4 и PAF.
Спектр цитокинов, выделяемых эозинофилами и базофилами, сходен с таковым тучных клеток и включает типичные ^2-цитокины (рис. 4.33). Среди них патогенетическую роль в реализации отложенной фазы аллергических реакций играют 4 вышеупомянутых цитокина — IL-4, IL-5, IL-9 и IL-13. Их эффекты сильно перекрываются, однако между ними есть и определенное «разделение труда». Так, IL-4, играющий ключевую роль в подготовке аллергических процессов (синтез IgE, развитие тучных клеток), на этапе поздних проявлений играет меньшую роль, участвуя в ремоделировании слизистых оболочек. IL-5 служит главным фактором привлечения эозинофилов. На этапе поздних проявлений аллергии основная роль принадлежит IL-13, который обусловливает ремоделирование слизистых оболочек, усиливает секрецию слизи, поддерживает повышенную активность гладких мышц бронхов. IL-9 выступает в качестве вспомогательного фактора в большинстве упомянутых процессов.
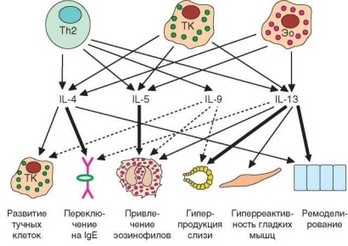
Рис. 4.33. Ключевые цитокины аллергических процессов, их клетки-продуценты и клетки-мишени. Цитокины, секретируемые эффекторными клетками аллергии, отвечают за ряд ключевых проявлений аллергии и связанных с ней заболеваний. Th2 — ^2-клетки; ТК — тучные клетки; Эо — эозинофилы
IL-5, IL-4 и IL-13, а также гуморальные факторы и белки из гранул эозинофилов определяют развитие местной воспалительной реакции по типу «эозинофильного воспаления», патогенетически отличающегося от классического «макрофагального» воспаления. Этот тип воспаления лежит в основе аллергических заболеваний — бронхиальной астмы, аллергического ринита, атопического дерматита. Общим индуцирующим фактором для обоих типов воспаления служит фактор некроза опухоли a (TNFa), секретируемый при аллергии ^2-лимфоцитами и тучными клетками.