
Биологическая фиксация молекулярного азота - одна из самых важных проблем почвенной микробиологии. Разрешение ее имеет большое практическое и теоретическое значение.
Азот - это важнейший биогенный элемент, входящий в состав белковой молекулы каждого живого существа. Его источником служит минеральный азот атмосферы и почвы. Но запасы минерального азота в почве сравнительно невелики - до 150 кг на гектар пахотного слоя, в то время как в атмосфере количество свободного (молекулярного) азота неисчерпаемо: около 4/5
окружающего нас воздуха приходится на долю свободного азота. Столб воздуха над гектаром почвы содержит его до 80000 т. Однако в силу своей инертности и ограниченной способности вступать в химические реакции с другими элементами этот азот непригоден для питания растений и животных. Именно поэтому он и получил свое название «азот», что означает «безжизненный».
Растения, выращиваемые на почвах, бедных минеральным азотом, отличаются слабым ростом и быстро гибнут, несмотря на наличие огромного количества молекулярного азота в воздухе. Этот азот оставался бы мертвым капиталом природы, если бы не микроорганизмы. Специфическая группа микроорганизмов, получившая название азотфиксаторов, обладает способностью фик
сировать атмосферный азот и переводить его в связанное состояние. За счет деятельности азотфиксаторов в почву ежегодно поступает 60-75 % азота от общего содержания его в почве. Согласно подсчетам, только культурные растения земного шара за год потребляют около 100 млн тонн азота, в то время как в виде минеральных удобрений в почву вносится приблизительного лишь 32 млн тонн, а остальной азотный дефицит покрывается за счет биологической азотфиксации.
Изучение природы биологической фиксации азота начато С. Н. Виноградским в 1893 г. и продолжено В. Л. Омелянским, С. П. Костычевым, М. В. Федоровым, Е. Н. Мишустиным и многими зарубежными учеными. В результате многочисленных исследований установлено, что способностью фиксировать молекулярный азот обладают не единичные виды, как это предполагалось до 1949 г., а многие микроорганизмы, принадлежащие к различным группам прокариот.
Азотфиксирующие микроорганизмы можно разделить на две группы: свободноживущие и симбиотические, т. е. вступающие в сожительство с высшими растениями.
Свободноживущие азотфиксаторы. Классическими представителями этой группы микроорганизмов являются бактерии рода Azotobacter и анаэробная палочка Cl. pasteurianum. Азотобактер был выделен в чистую культуру голландским ученым М. Бейеринком в 1901 г., который описал его морфологию и физиологию. В молодой культуре азотобактер представляет собой крупные (3x6 мкм), иногда сцепленные попарно палочки с закругленными концами. С возрастом они постепенно укорачиваются и превращаются в кокки, покрытые толстой слизистой капсулой (рис. 12.2). Кокки * часто бывают соединены в сарциноподобные пакеты - цисты, характерные для покоящейся стадии азотобактера.
Молодые клетки азотобактера благодаря расположенным по всей поверхности тела жгутикам обладают подвижностью. Последняя при старении клетки в результате уменьшения числа жгутиков и появления капсулы исчезает.
Азотобактер хорошо растет на безазотистых питательных средах. На плотных средах колонии имеют вид густослизистых полупрозрачных выпуклых капель. В зависимости от вида азотобактера они окрашиваются в желтый (Az. vinelandii), темно- коричневый (Az. chroococcum) или флуоресцирующий желто- зеленый цвет (Az. agile).
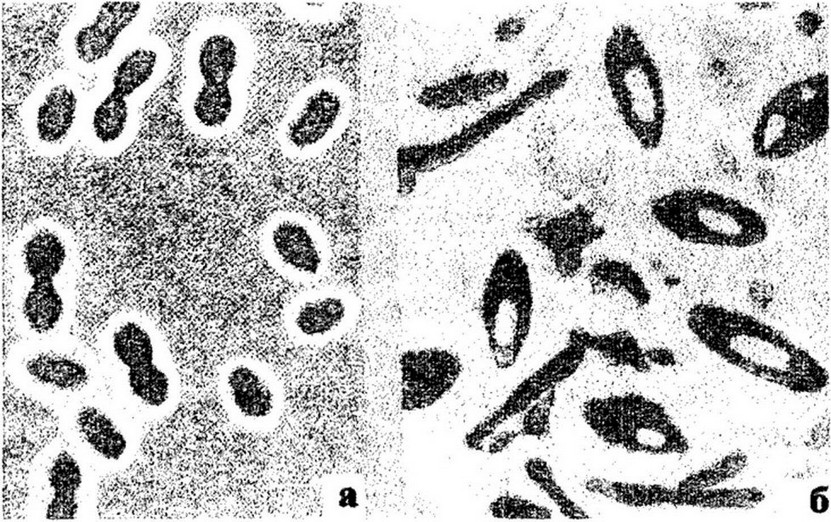
Рис. 12.2. Азотфиксирующие бактерии: а - Az. chroococcum (увеличение х 1 800); б - Cl. pasteurianum (увеличение хЗ 500)
Углерод и энергию азотобактер получает из разнообразных органических веществ: углеводов, спиртов, органических кислот и их солей. Источником азота, кроме атмосферного, могут служить минеральные (соли аммония, нитраты и нитриты) и органические (аминокислоты, мочевина) соединения. Но в тех случаях, когда азотобактер развивается на азотсодержащих средах, он не фиксирует молекулярный азот и при длительном культивировании на таких средах вообще утрачивает азотфиксирующую способность. Количество азота, фиксированное на 1 г потребленного источника энергии, называется азотфиксирующей способностью. Эта величина непостоянна, она зависит от индивидуальных свойств штамма, состава питательной среды, ее кислотности, условий выращивания культуры (температура, аэрация) и в среднем равна 10-15 мг.
Азотобактер - строгий аэроб. Оптимальная температура для него 28-30° С, pH 7,2-8,0. Хотя азотобактер в естественных условиях встречается при pH от 4,5 до 9,0, как в кислых, так и в сильно щелочных средах, он теряет активность и способность связывать молекулярный азот. Азотобактер требователен к наличию в среде фосфора, кальция и микроэлементов - молибдена, железа, кобальта, в первую очередь необходимых для осуществления процесса азотфиксации.
Азотобактер обладает способностью синтезировать значительное количество биологически активных веществ - витамины группы В, никотиновую и пантотеновую кислоты, гетероауксин, гиббереллин и фунгистатический (задерживающий рост грибов) антибиотик группы анисомицина.
Азотобактер широко распространен в почвах и водоемах, имеющих нейтральную или слабощелочную реакцию среды и содержащих легкодоступные органические вещества.
Морфологическое сходство с азотобактером имеют аэробные азотфиксирующие бактерии рода Beijerinckia. Клетки имеют палочковидную, круглую или овальную формы. На безазотистых плотных средах они образуют выпуклые, иногда складчатые слизистые колонии. Продуктивность азотфиксации 15, 20 мг азота на 1 г сахара. От азотобактера эти бактерии отличаются медленным ростом, высокой кислотоустойчивостью (развиваются при pH 3,0), отрицательным отношением к наличию в среде кальция. Бактерии рода Beijerinckia широко распространены в южной и тропической зонах, реже встречаются в зоне умеренного климата.
К свободноживутцим азотфиксаторам относится и анаэробный микроорганизм Cl. pasteurianum, клетки которого имеют вид палочки длиной 1,5-8 мкм, шириной 0,8-1,3 мкм с округленными концами (см. рис. 12.2). При образовании спор клетки утолщаются посередине и принимают форму веретена. Эти бактерии - грампо- ложительные облигатные анаэробы, предельное pH для них равно 5,5-8,0; споры выдерживают температуру 96° С, а при 100° С погибают. Подобно азотобактеру данные микроорганизмы используют различные источники углеродного питания. На безазотистых средах они фиксируют молекулярный азот.
Cl. pasteurianum широко распространен во всех почвах. Особенно в больших количествах он встречается в плохо аэрируемых слежавшихся или залитых водой почвах. Рисовые поля обогащаются азотом в основном за счет деятельности Cl. pasteurianum и
водорослей и длительное время не нуждаются в азотных удобрениях.
Связывать молекулярный азот воздуха могут и некоторые другие микроорганизмы - Azotomonas fluorescens, Azotomonas insolita, микобакетрии, фотосинтезирующие бактерии, а также азоспириллы, обитающие в ризосфере злаковых растений. Однако азот- фиксирующая активность у этих микроорганизмов гораздо ниже, чем у первых двух групп.
Симбиотические азотфиксаторы. Наряду со свободноживу- щими азотфиксирующими микроорганизмами в почве обитают и симбиотические азотфиксаторы - клубеньковые бактерии, открытые русским ученым М. С. Ворониным в 1866 г. Воронин обнаружил на корнях люпина и ольхи мелкие клубеньки, заполненные палочковидными организмами вздутой или ветвистой формы, описал морфологию и размеры этих организмов, считая их причиной образования клубеньков.
Через 20 лет это открытие было подтверждено М. Бейеринком. Более того, Г. Гельригелем и Г. Вильфартом было доказано, что бактерии, находящиеся в клубеньках, выполняют важную биологическую функцию фиксации молекулярного азота. (Данная гипотеза была высказана еще в 1838 г. Ж. Буссенго). Г. Гельригель и Г. Виль-

 фарт установили, что клубеньки образуются только на корнях бобовых растений и только в нестерильной почве. Отмечено, что в этой же почве накапливается значительно больше азота, чем его было до выращивания бобовых растений. Так была доказана роль клубеньков бобовых растений в ассимиляции атмосферного азота. Чистая культура клубеньковых бактерий, выделенная в 1888 г. М. Бейеринком из клубеньков гороха, была названа им Bacillus radicicola. Затем было предложено название Rhizobium ( от греч. rtiizo - корень, bios - жизнь, т. е. жизнь на корнях). В настоящее время эти
фарт установили, что клубеньки образуются только на корнях бобовых растений и только в нестерильной почве. Отмечено, что в этой же почве накапливается значительно больше азота, чем его было до выращивания бобовых растений. Так была доказана роль клубеньков бобовых растений в ассимиляции атмосферного азота. Чистая культура клубеньковых бактерий, выделенная в 1888 г. М. Бейеринком из клубеньков гороха, была названа им Bacillus radicicola. Затем было предложено название Rhizobium ( от греч. rtiizo - корень, bios - жизнь, т. е. жизнь на корнях). В настоящее время этибактерии объединены в самостоятельный род Rhizobium.
Клубеньковые бактерии представляют собой грамотрица- тельные, не образующие спор, подвижные палочки размером 1,2-3 х 0,5-0,9 мкм (рис. 12.3). В клубеньках клетки обычно более крупных размеров, часто имеют раздутую или ветвистую формы, напоминающие буквы V, L, X. Такие формы принято называть бактероидами.
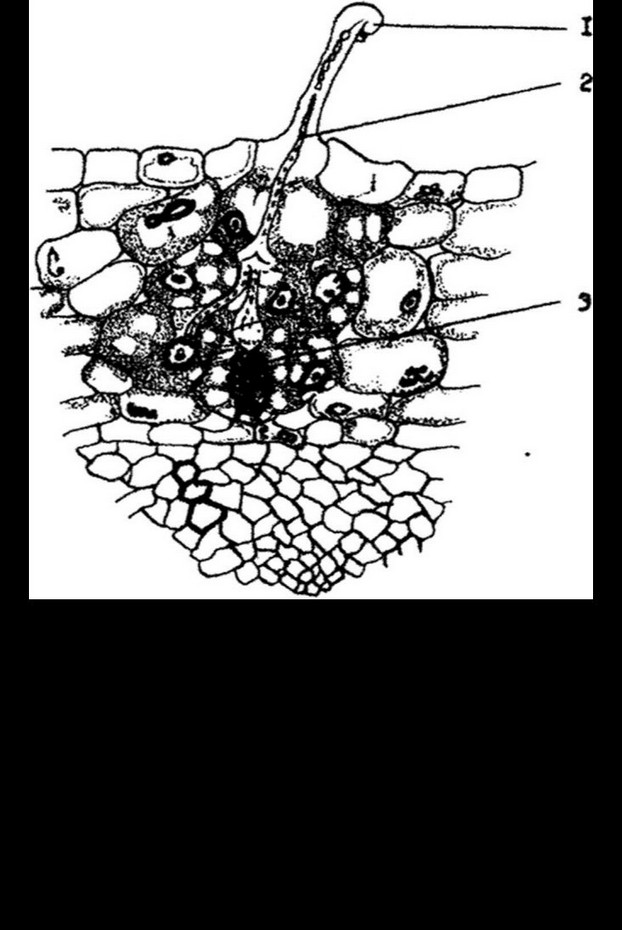 Клубеньковые бактерии чаще всего обнаруживаются там, где растут бобовые растения, хотя могут встречаться в почве и в свободном состоянии. В корни растений они проникают через корневые волоски, где образуют инфекционные нити, которые представляют собой бактерии, заключенные в выделяемую ими слизь (рис. 12.4). Эти нити проникают через стенки клеткок до внутренних покровов коры корня. Под действием р-индол-уксусной кислоты или других каких-то факторов, связанных с инфекцией, клетки коры начинают беспорядочно делиться и в результате возникает первичная, затем вторичная меристемы клубенька. Инфекционная нить, разветвляясь, пронизывает клетки меристемы и бактерии под действием внутреннего давления, возникающего вследствие их интенсивного размножения, выходят из нити в цитоплазму клеток растения-хозяина. Здесь они продолжают размножаться, а клетки растений, наполняющиеся бактериями, начинают усиленно делиться. Так происходит формирование клубенька (рис. 12.5). Зона клубенька, заполненная бактериями, называется
Клубеньковые бактерии чаще всего обнаруживаются там, где растут бобовые растения, хотя могут встречаться в почве и в свободном состоянии. В корни растений они проникают через корневые волоски, где образуют инфекционные нити, которые представляют собой бактерии, заключенные в выделяемую ими слизь (рис. 12.4). Эти нити проникают через стенки клеткок до внутренних покровов коры корня. Под действием р-индол-уксусной кислоты или других каких-то факторов, связанных с инфекцией, клетки коры начинают беспорядочно делиться и в результате возникает первичная, затем вторичная меристемы клубенька. Инфекционная нить, разветвляясь, пронизывает клетки меристемы и бактерии под действием внутреннего давления, возникающего вследствие их интенсивного размножения, выходят из нити в цитоплазму клеток растения-хозяина. Здесь они продолжают размножаться, а клетки растений, наполняющиеся бактериями, начинают усиленно делиться. Так происходит формирование клубенька (рис. 12.5). Зона клубенька, заполненная бактериями, называется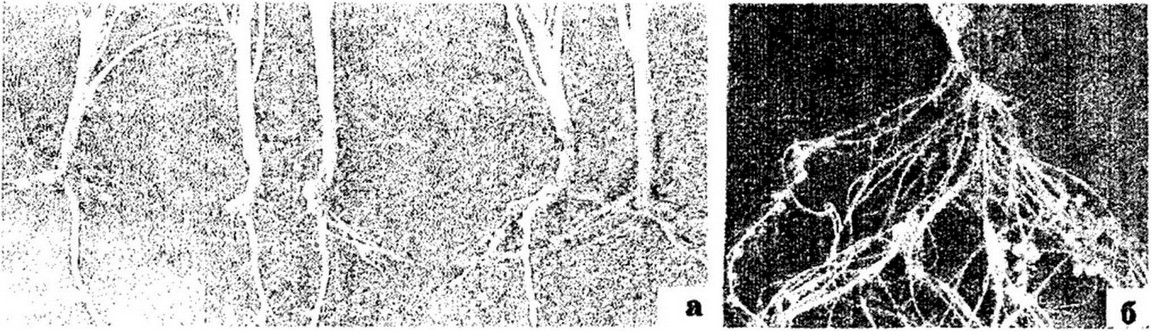
Рис. 12.5. Клубеньки на корнях люпина (а) и гороха (б)
бактериальной зоной. Она занимает центральную часть клубенька, окружена меристемой, покрытой клубеньковой корой. Ни клетки меристемы, ни клетки коры не содержат бактерий.
Связь клубенька с корнем растения осуществляется через сосудисто-волокнистые пучки, которые образуются из клеток перикамбия центрального цилиндра корня. По этим пучкам в клубеньки поступают питательные вещества, синтезируемые растениями, и выводятся продукты жизнедеятельности бактерий. Так между бактериями и растениями устанавливаются тесные симбиотические отношения.
Клубеньковые бактерии характеризуются строгой специфичностью в отношении растений. Они могут вступать в симбиоз только с определенным видом бобового растения.
Специфичность клубеньковых бактерий определяется природой капсульных липополисахаридов, которые избирательно реагируют с лектинами (фитогемагглютининами) бобовых растений. Последние представляют собой белки, продуцируются бобовыми растениями, концентрируясь на поверхности корней. Они несут ответственность за «узнавание» специфических клубеньковых бактерий. Взаимодействие с бактериями осуществляется аналогично взаимодействию антиген-антитело. Лектины играют роль антигена, липополисахариды капсулы бактерий - роль антитела. В результате их взаимосвязи клубеньковые бактерии фиксируются на поверхности корней и инфицируют растение. Мутанты бактерий с нарушенной способностью к синтезу липополисахаридов неспособны заражать растения и образовывать клубеньки.
Поскольку лектин каждого вида бобового растения связывается только с определенным липополисахаридом Rhizobium, образуется видоспецифичный комплекс, присущий конкретному виду бактерий и растений. Специфичность клубеньковых бактерий положена в основу их классификации.
Различают бактерии клевера Rh. trifolii, бактерии фасоли Rh. phaseoli, бактерии люпина Rh. lupini, бактерии сои Rh. japonicum, бактерии люцерны и донника Rh. meliloti, бактерии гороха, вики, бобов Rh. ieguminosarum.
При инфекции корневой системы бобовых растений клубеньковыми бактериями большое значение имеет вирулентность последних. Под вирулентностью следует понимать способность бактерий проникать в растение и вести там активный образ жизни, т. е. размножаться и вызывать образование клубеньков. Если специфичность клубеньковых бактерий определяет спектр их действия, то вирулентность характеризует их активность в пределах данного спектра.
Однако не все бобовые растения способны образовывать клубеньки. Из изученных 1 200 видов 133 не формируют их. Есть предположение, что причиной является отсутствие бактерий, специфичных для этих видов растений.
Среди клубеньковых бактерий различают активные и неактивные расы, которые образуют разные по эффективности клубеньки. Эффективные клубеньки характеризуются более крупными размерами и интенсивной азотфиксацией. Неэффективные, как правило, лишены азотфиксирующей способности. Причина эффективности азотфиксации долгое время оставалась неизвестной. К выяснению ее подходили по-разному: рассматривались морфофизиологические, биохимические и генетические особенности бактерий. Однако вскоре пришли к убеждению, что, основываясь лишь на свойствах бактерий, разрешить этот вопрос невозможно, так как оказалось, что ни культуральная, ни физиологическая специфика, ни антигенная структура их не связаны с эффективностью клубеньков. И только изучение внутреннего содержимого клубенька в бактериологическом и биохимическом отношении позволило установить, что эффективные клубеньки заполнены бактериями в стадии бактероидов. Второй особенностью эффективных клубеньков является наличие в них гемоглобина, который называют леггемоглобином (от «гемоглобин»
Leguminosae»). Он был выделен из клубеньков в 1939 г. японским ученым Г. Кубо. Данное открытие имело большое значение, так как позволило установить химические различия между эффективными и неэффективными клубеньками.
Леггемоглобин рассматривается как необходимый фактор симбиотической фиксации азота, где он выполняет функции регулятора парциального давления кислорода и транспорта электронов в азотфиксирующую систему. Предполагают также, что леггемоглобин может осуществлять превращения гидроксиламина в аммиак, азот которого включается в состав органических веществ.
В период максимальной азотфиксации, обычно наблюдающейся в стадии бутонизации и цветения растений, в них поступает до 90 % азота, фиксированного клубеньками. Установлено, что бобовые растения в симбиозе с эффективными штаммами клубеньковых бактерий фиксируют от 50 до 200 кг азота на 1 га почвы.
Кроме бобовых растений способностью к образованию клубеньков и симбиотической азотфиксации обладает более 200 видов небобовых. Среди них представители голосеменных (саговники, гинкговые, хвойные) и покрытосеменных (береза, ольха, облепиха). Из травянистых небобовых растений клубеньки обнаружены у дикорастущих злаковых (лисохвост луговой, мятлик луговой, два вида волоснеца - сибирский и солончаковый).
В клубеньках небобовых растений выявлены грибы фико- мицеты у хвойных, актиномицеты у ольхи, бактерии у злаковых. Все перечисленные представители способны к симбиотической азотфиксации. Например, роща ольхи (в среднем 5 растений на 1 м2) за 7 лет дает прибавку азота 700 кг на 1 га.
Многие тропические растения (около 400 видов) образуют клубеньки на листьях, так называемые листовые клубеньки. Они располагаются на нижней поверхности листа. Так, возбудителями формирования листовых клубеньков у растений Pavetta и Psychotria являются бактерии рода Klebsiella. Эти бактерии фиксируют азот как в симбиозе с растениями, так и в чистой культуре. Полагают, что они обеспечивают развитие растений на почвах, бедных азотом.