
Бактерии подобно высшим организмам обладают способностью к обмену генетического материала, но существенно отличаются от последних способами передачи его от донорной клетки к реципиентной. Обмен генетического материала у бактерий имеет место при трансформации, конъюгации и трансдукции. Исторически у бактерий раньше других описано явление трансформации.
Трансформация (превращение, перестройка) - это изменение генома бактерии-реципиента под влиянием поглощенной из среды свободной ДНК, выделенной из бактерии-донора. Трансформацию может осуществлять только ДНК, включающаяся в геном реципиента. Ее источником могут быть свежеубитые бактерии или чистые препараты, экстрагированные из бактерий. Поэтому для
получения трансформантов реципиентные бактерии выращивают на средах, содержащих чистую ДНК или убитые клетки доноров.
Трансформация была открыта в 1928 г. английским ученым Ф. Гриффитсом. Он установил превращение бескапсульного пневмококка R-типа в капсульный вирулентный S-тип. Заражая мышей смешанной взвесью живых бескапсульных авирулентных пневмококков и убитых нагреванием капсульных вирулентных пневмококков, Ф. Гриффитс наблюдал гибель животных, из крови которых выделял наряду с бескапсульными и капсульные пневмококки. Автор пришел к заключению, что бескапсульные варианты приобрели способность образовывать капсулу под влиянием капсульных пневмококков, несмотря на то, что последние были мертвыми. Однако природу трансфер мир ующего вещества Ф. Гриффитс не установил. Он считал, что ответственными за образование капсулы являются полисахариды капсульных пневмококков S-типа.
Опыты Ф. Гриффитса по трансформации капсулы у пневмококков подтвердились исследователями. Так, в 1931 г. М. Даусон и Р. Сиа описали аналогичную трансформацию in vitro, выращивая клетки R-типа в бульоне, содержащем клетки пневмококков S-типа, убитые нагреванием. Позднее С. Алловий (1933) показал, что такие же превращения R-типа в S-тип можно получить, использовав бесклеточные экстракты капсульных пневмококков.
Хотя трансформацию бескапсульных пневмококков наблюдали многие исследователи, природа трансформирующего агента оставалась неизвестной. И только в 1944 г. О. Эвери с сотрудниками выделил трансформирующее вещество из капсульных убитых пневмококков и исследовал его свойства. Оно оказалось чувствительным к ДНК-азе. Применение высокоочищенного препарата ДНК-азы показал о, что он подавляет активность трансформирующего вещества. Это явилось неоспоримым фактом того, что трансформацию вызывает ДНК. Дополнительные доказательства были получены в опытах с использованием ДНК, максимально очищенной от примесей белка, полисахаридов и других клеточных компонентов. Эта ДНК проявила высокую активность: в разведении 1: 600 млн она способна была вызвать специфическую трансформацию.
Установление трансформирующей роли ДНК у щим аргументом в пользу того, что генетическая инс дится в ДНК, а не в белке, как предполагали.
Возможность передачи признаков посредством ной к реципиентной клетке была показана и на других видах бактерий и бацилл: Rhizobium, Neisseria, Вас. subtilis.
Трансформирующей активностью обладает лишь высокомолекулярная (М gt; 105) двухцепочечная ДНК, хотя в геном реципиента включается только одна цепь, а другая разрушается ДНК-а зой реципиентной клетки.
Бактериальные клетки различаются по способности к трансформации. Это зависит от их генетических способностей: от выделения ДНК-азы, образования капсулы, препятствующей проникновению ДНК, от состава среды, влияющей в свою очередь на физиологическое состояние клеток, и др.
Оптимальное физиологическое состояние клеток, в котором они способны к поглощению чужеродной ДНК, называется компетентностью. Компетентность обусловлена появлением на поверхности клетки особого антигена - фактора компетентности, который является низкомолекулярным белком и играет роль специфического ДНК-связывающего рецептора. Инициируют компетентность три белка: автолизин, ДНК-связывающий белок и эндонуклеаза I. В период развития компетентности происходят изменения структуры клеточной стенки, в результате чего стенка становится более пористой и проницаемой для ДНК.
Процесс трансформации разделяют на несколько стадий: 1) присоединение ДНК к поверхностным рецепторам реципиентной клетки; 2) проникновение ДНК в клетку; 3) превращение проникшей двухцепочечной ДНК в одноцепочечную; 4) рекомбинация проникшей ДНК с ДНК реципиента; 5) фенотипическое выражение поглощенного гена (или генов). На первой стадии трансформирующая двухцепочечная ДНК связывается с поверхностным рецептором компетентной клетки. В этот период она чувствительна к ДНК-азе. Затем, сразу после присоединения перед проникновением в клетку эндонуклеазы, находящиеся в периплазматическом пространстве клетки надрезают поочередно обе цепи ДНК с образованием более коротких фрагментов двухцепочечной ДНК (5 ' 106 - Г 107 п. н ).
Эти фрагменты подвергаются действию экзонуклеаз, которые разрушают одну цепь ДНК. В клетку проникает фрагмент одноцепочечной ДНК. Для проникновения используется энергия
трансмембранного потенциала
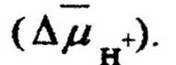 Далее следует рекомбинация одноцепочечного фрагмента с двухцепочечной ДНК реципиента и образование трансформантов (рекомбинантов). Так как при трансформации передается небольшой фрагмент ДНК донора, то образующиеся трансформанты обычно обладают одним новым признаком. Иногда могут трансформироваться и два признака. Это имеет место в тех случаях, когда поглощенный фрагмент ДНК содержит тесно сцепленные гены или клетка трансформирована двумя фрагментами ДНК-донор а. Частота трансформации более высока, если донором и реципиентом являются бактерии одного вида. Трансформация может иметь место и между бактериями разных видов, но с очень низкой частотой образования трансформантов. Так, частота трансформантов Haemophilus influensae, устойчивых к стрептомицину, при трансформировании бактерий ДНК, полученной из клеток этого же вида, составляет 1 ’ 10*, при трансформировании бактерий другого вида (Н. suis) выход рекомбинантов в 104 раз ниже.
Далее следует рекомбинация одноцепочечного фрагмента с двухцепочечной ДНК реципиента и образование трансформантов (рекомбинантов). Так как при трансформации передается небольшой фрагмент ДНК донора, то образующиеся трансформанты обычно обладают одним новым признаком. Иногда могут трансформироваться и два признака. Это имеет место в тех случаях, когда поглощенный фрагмент ДНК содержит тесно сцепленные гены или клетка трансформирована двумя фрагментами ДНК-донор а. Частота трансформации более высока, если донором и реципиентом являются бактерии одного вида. Трансформация может иметь место и между бактериями разных видов, но с очень низкой частотой образования трансформантов. Так, частота трансформантов Haemophilus influensae, устойчивых к стрептомицину, при трансформировании бактерий ДНК, полученной из клеток этого же вида, составляет 1 ’ 10*, при трансформировании бактерий другого вида (Н. suis) выход рекомбинантов в 104 раз ниже.
Путем трансформации передаются разные свойства: образование капсул, устойчивость к антибиотикам, способность к синтезу аминокислот, витаминов и др.
По своей природе к трансформации близка трансфекция. Это перенос генетической информации фага компетентным клеткам бактерий посредством ДНК, выделенной из частиц фага. В таких клетках, инфицированных фаговой ДНК, происходит развитие нормальных, полноценных фагов. Трансфекцию удалось осуществлять не только с помощью ДНК фагов, но и ДНК вирусов животных.
- Конъюгация
Конъюгация (спаривание) - передача генетического материала от донорной к реципиентной клетке при их непосредственном контакте. Конъюгация осуществляется только между клетками разного пола. Пол у бактерий определяется наличием или отсутствием полового фактора - F-фактора, который представляет собой кольцевую молекулу ДНК и относится к категории плазмид. Бактерии, которые содержат F-фактор, являются бактериями мужского типа и служат донорами генетического материала. Другие бактерии не имеют F-фактора - это бактерии женского типа. Они являются реципиентами генетического материала, передаваемого донорными штаммами.
Конъюгация прокариот является аналогом полового процесса эукариот.
Открытие конъюгации бактерий принадлежит Дж Ледербергу и Е. Татуму (1946). Они использовали два ауксотрофных мутанта Е. coli К-12, каждый из которых в отдельности не обладал способностью синтезировать две аминокислоты. Один был ауксотрофным по аминокислотам Л и Л, но синтезировал кислоты С и D (А~В~С+D+), другой мутант был комплементарен (А~B C D). На минимальной среде эти мутанты раздельно не росли. При высеве смеси их на ту среду появлялись колонии, клетки которых обладали способностью синтезировать все 4 аминокислоты, т. е. это были генетические рекомбинанты двух реципрокно дефектных (взаимодополняющих) родительских клеток. Однако в этом опыте не исключалась возможность появления рекомбинантного потомства под влиянием веществ, обладающих трансформирующей активностью.
Наиболее убедительные доказательства образования генетических рекомбинантов в результате конъюгации были получены Б. Дэвисом. В одно колено ?/-образной трубки, разделенной стеклянным пористым бактериальным фильтром, помещался один ауксотрофный штамм бактерий, в другое - другой. Наличие пористого фильтра исключало физический контакт бактерий, но не препятствовало диффузии трансформирующих веществ из одного колена в другое. Спустя некоторое время из содержимого каждого колена производился высев бактерий на минимальную среду, однако ни в одном из них прототрофов не было обнаружено, т. е. рекомбинанты не образовывались. Когда же оба родительских штамма засевали в одно и то же колено трубки, что позволяло клеткам вступать в прямой контакт, рекомбинанты появлялись.
Наличие такого контакта между клетками удалось наблюдать в 1957 г. непосредственно с помощью электронного микроскопа (рис.
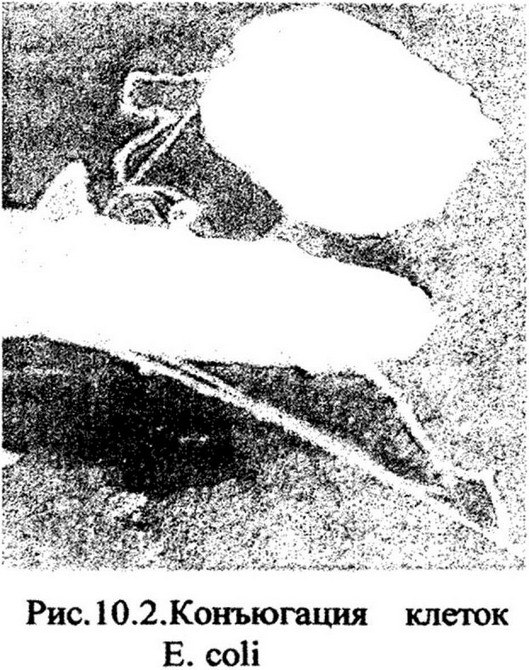 10.2). Позже было установлено, что конъюгирующие клетки соединяются через конъюгационный мостик, образованный половой ворсинкой F-пили донорной клетки.
10.2). Позже было установлено, что конъюгирующие клетки соединяются через конъюгационный мостик, образованный половой ворсинкой F-пили донорной клетки.
Сближению клеток способствует сокращение половой ворсинки или втягивание ее внутрь донорной клетки.
Реципиентная клетка подтягивается к донорной до непосредственного контакта клеточных стенок. Репликация ДНК осуществляется по механизму «катящегося кольца» (рис. 10.3). Когда клетки вступили в контакт, в молекуле ДНК донорной клетки происходит одноцепочечный надрез, что создает условия для раскручивания двойной цепи. Одна из цепей,
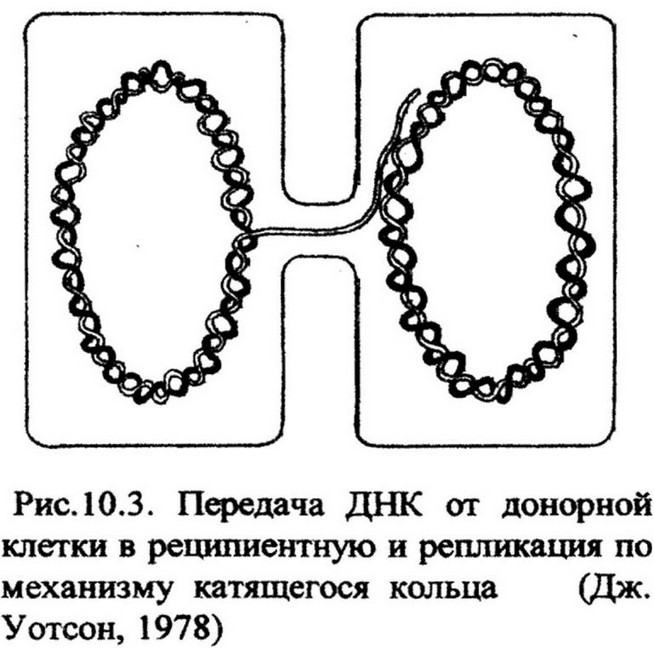 начиная с 5'-конца, передается в реципиентную клетку, другая остается в донорной. Одновременно с передачей происходит и репликация ДНК. По мере передачи и раскручивания цепи ферменты ДНК-по- лимеразы в донорной и реципиентной клетках одновременно синтезируют комплементарные цепи. Таким образом, от донора к реципиенту передается одна цепь, но в процессе передачи она
начиная с 5'-конца, передается в реципиентную клетку, другая остается в донорной. Одновременно с передачей происходит и репликация ДНК. По мере передачи и раскручивания цепи ферменты ДНК-по- лимеразы в донорной и реципиентной клетках одновременно синтезируют комплементарные цепи. Таким образом, от донора к реципиенту передается одна цепь, но в процессе передачи она
достраивается и ковалентно замыкается в кольцевую структуру. Затем путем генетической рекомбинации она включается в хромосому реципиента, реплицируется и в результате деления клетки образуется рекомбинантное потомство. Анализ рекомбинантов показал, что клетка-донор передает реципиентной клетке лишь часть своего генома и только в редких случаях возможна передача его целиком. В связи с этим образуется мерозигота. Она содержит полный геном реципиента и лишь фрагмент генома донора.
Конъюгация - однонаправленный, или ассиметричный процесс, т. е. перенос генетического материала происходит в одном направлении - от донорной (мужской) клетки к реципиентной (женской).