
В середине 70-х г. XX в. открыты подвижные генетические элементы. Они представляют собой сегменты ДНК, способные к транспозиции (перемещению) в пределах одного либо разных геномов. По степени сложности строения различают три типа мигрирующих генетических элементов: IS-элементы (от англ, insertion sequence - вставочные последовательности), транспозоны (Tn-элементы) и некоторые бактериофаги, в частности фаг Мю.
Простейшими генетическими структурами, способными к транспозиции, являются IS-элементы. Размер их составляет в среднем 750-1500 пар нуклеотидов (п. н.). Они содержат только гены, обеспечивающие их собственное перемещение. В структуре IS-элементов различают центральную часть и ограничивающие (фланкирующие) концевые повторы. В центральной части расположены гены, кодирующие синтез белков, необходимых для транспозиции. Концевые участки представлены повторяющимися нуклеотидными последовательностями, длиной 8-40 п. н. Повторения имеют противоположную друг другу ориентацию и называются инвертированными (перевернутыми) повторами. Они служат отличительным признаком различных мигрирующих генетических элементов.
Структура концевых повторов определяет размеры дупликаций (удвоение) ДНК в местах внедрения IS-элементов. Так, IS 1-элемент, обнаруженный в составе хромосомы Е. coli-K12, состоит из 768 п. н., образующих на концах инвертированные повторы длиной по 30 п. н. каждый. Всякий IS-элемент имеет свою нуклеотидную последовательность и может в любой ориентации включаться в ДНК бактерий, плазмид и фагов, вызывая инактивацию отдельных структурных генов и, как следствие, мутации генома или нарушая регуляторные функции оперона. В бактериальной хромосоме может содержаться одновременно несколько копий одного и того же IS-элемента. Перемещение IS- элементов индуцирует разные виды хромосомных перестроек - дупликации, инверсии, делеции.
Транспозоны, или Tn-элементы - подвижные генетические элементы содержат гены фенотипических свойств бактерий и гены
собственной транспозиции. Они способны внедряться в разные участки хромосомы или во внехромосомные генетические структуры. Транспозоны отличаются от IS-элементов более сложной организацией, а некоторые содержат в своем составе IS-элементы.
Транспозоны разделяют на два класса: А и Б (рис. 10.4). Транспозоны класса А (Тп 5) в центральной части содержат структурные гены, которые детерминируют фенотипические свойства, например, устойчивость бактерий к антибиотикам, а гены транспозиции содержатся в составе концевых инвертированных повторов, которыми являются IS-элементы. У транспозонов класса Б (Тп 3) в центральной части содержатся не только гены фенотипических признаков, но и гены транспозиции. Концевые повторы их значительно короче и не могут выполнять функции транспозиции. Эти функции выполняют два гена центральной части. Различия между транспозонами класса А и класса Б состоят также в размерах образуемых дупликаций при внедрении их в плазмиды или хромосомы: первые образуют дупликации 9 нуклеотидных пар, вторые - только 5.
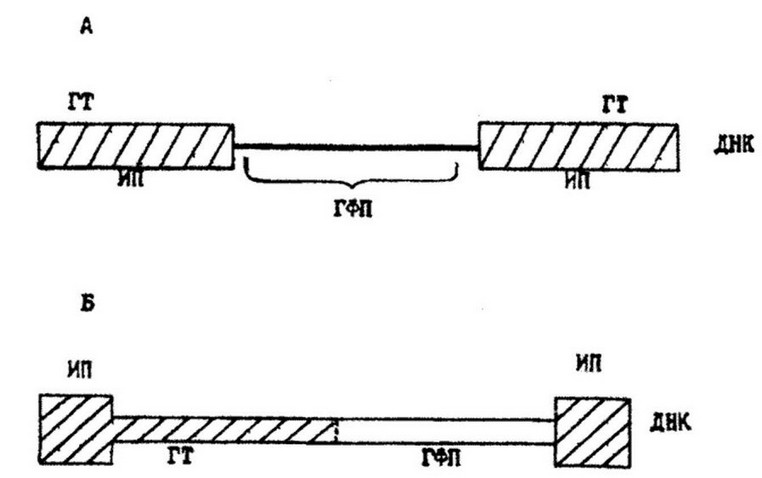
Рис. 10.4. Схема структуры транспозонов класса А и класса Б: ИП - инвертированные повторы; ГТ - гены транспозиций; ГФП - гены фенотипических признаков
Транспозоны имеют значительно большие размеры, чем IS- элементы и составляют в среднем 3 500-15 000 пар нуклеотидов. Так, общая протяженность транспозона Тп 5 составляет 5 800 п. н., из них по 1 500 п. н. приходится на инвертированные концевые повторы. Тп 5 кодирует пять белков. Из них один белок кодирует центральная часть и по два белка - концевые повторы. Транспозон Тп 5 детерминирует устойчивость к канамицину, неомицину и другим родственным антибиотикам.
Следствием перемещения транспозонов, как и IS-элементов, могут быть различные хромосомные перестройки: делеции, инверсии, транслокации, дупликации. Помимо них, перемещение транспозонов между двумя различными репликонами (двумя плазмидами, или плазмидой и хромосомой) может вызывать слияние этих репликонов с образованием коинтегратов. Последующая сайт-специфи- ческая рекомбинация приводит к разделению коинтеграта на два репликона с включением в каждый репликон по одной копии транс- позона. Регуляция транспозиции осуществляется собственными генами МГЭ и хромосомными генами бактерий-хозяев.
Умеренный фаг Мю, выделенный в 1963 г. из культуры холерного вибриона, также обладает свойствами МГЭ. Однако, в отличие от IS-элементов и транспозонов он не содержит на концах генома ни прямых, ни инвертированных нуклеотидных последовательностей. Концевые повторы фага Мю составляют фрагменты ДНК клетки-хозяина, в которой развивался фаг. ДНК клетки прикрепляется к геному фага при его размножении и теряется в ходе его интеграции в новый сайт. Уникальной способностью фага Мю является перенос генов бактерий в различные участки хромосомы или плазмиды клетки-реципиента. Фаг Мю осуществляет постоянную транспозицию в ходе всего литического цикла. Он не обладает специфичностью в отношении локуса хромосомы и может спонтанно внедряться в разные участки вдоль всей хромосомы, вызывая мутации хромосомных генов. За высокую активность индуцировать мутации он получил название Мю (от англ, mutator).
Несмотря на некоторые различия в структурной организации, общим свойством МГЭ является их способность внедряться во множество участков хромосомной или плазмидной ДНК, вызывая мутации и различные генные перестройки. МГЭ служат также специфическими сайтами внедрения плазмид в хромосомы. Через посредство МГЭ осуществляется рекомбинация между негомологичными ДНК. Временную область гомологии создают МГЭ,
включаясь в тот или иной участок хромосомной или плазмидной
ДНК.
Мигрирующие генетические элементы, индуцируя генные и хромосомные перестройки, вносят существенный вклад в перераспределение генетической информации, обеспечивают бактериям селективные преимущества в определенных условиях существования, оказывают существенное влияние на развитие и эволюцию микробных видов.