АПОПТОЗ
Термин "апоптоз" появился в литературе в 1972 г. для обозначения особого типа гибели' клеток, отличного от некроза. Установлено, что в клетках ' организма существует генетическая программа, которая обеспечивает определенный по времени жизненный цикл и при определенных физиологических или патологических условиях включает программу гибели клетки.
Таким образом, апоптоз, или программированная (физиологическая) смерть клетки, является тем механизмом, посредством которого осуществляется генетическая программа клеточной гибели. Морфологически апоптоз характеризуется конденсацией хроматина, фрагментацией ДНК и изменением мембраны клетки. В конце концов клетка фрагментируется и подвергается фагоцитированию без развития воспаления.
Апоптоз ' играет важную роль в следующих нормальных биологических процессах: эмбриональном развитии; регуляции состава и численности клеточных популяций в тканях взрослого организма (например в обновлении клеток иммунной системы); различного рода гормональных перестройках организма (например атрофия эндометрия у женщин в процессе менструального цикла).
Важна роль апоптоза и при различных патологических процессах. Наиболее полно она изучена при опухолевом росте. С одной стороны, развитие опухоли (гиперплазия в широком смысле) в некоторых случаях может быть следствием не усиления митоза, а ослаблением апоптоза. С другой стороны, усиление апоптоза имеет значение при регрессии опухолей — сегодня это одно из актуальных направлений лечения опухолей.
Таким образом, апоптоз является общим механизмом регулируемой (программированной) гибели клеток в ' нормальных и патологических процессах.
Какие же структуры со стороны ' иммунной системы и со стороны клетки-мишени участвуют во включении механизма апоптоза?
Одним из мембранных клеточных рецепторов, ответственных за контролируемый тканевой гомеостаз и иммунный ответ, является белок с молекулярной массой 45 kD, получивший название Fas-рецептора (CD95/APO-1). Он экспрессируется на клетках многих тканей, в том числе селезенки, лимфатических узлов, печени, легкого, почки, яичников и др.
Роль этого рецептора в "судьбе", в частности, гемопоэтических клеток чрезвычайно важна, поскольку связана со скоростью созревания и восстановления пула клеток. У мышей с отсутствием на мембране иммунных клеток Fas-рецептора развивались генерализованная лим- фаденопатия, лимфоцитоз, сплено- и гепатомегалия.
Кроме Fas-рецептора, на поверхности многих клеток, в том числе Т-лимфоцитов, нейтрофилов, макрофагов, тимоцитов идр., имеется еще один мембранный белок с молекулярной массой 4,0 • 104 D — Fas- лиганд (FASL). Он принадлежит к семейству опухольнекротизирующе- го фактора и впервые был обнаружен на мембране активированных Т- лимфоцитов. Впоследствии было установлено, что FasL может экспрессироваться на самых различных типах клеток (помимо указанных) в ответ на разнообразные стимулы, например на макрофагах, инфицированных ВИЧ-инфекцией, гепатоцитах, обработанных химиопрепаратами, а также на иных клетках при их опухолевой трансформации.
Fas-Лиганд имеет растворимую форму в виде белка с молекулярной массой 1,7 • 104 D.
Связывание Fas-рецептора с Fas-лигандом приводит к включению механизмов апоптоза. При этом мембранносвязанная форма FasL включает сигнал апоптоза при прямом контакте клетки с клеткой, тогда как растворимая форма FasL ответственна за киллинг клеток по типу аутокринного самоубийства или паракринной смерти близлежащей клетки.
Возвращаясь к специфическим Т-лимфоцитам-киллерам, следует подчеркнуть, что они реализуют свою киллинговую функцию, включая апоптоз, двумя независимыми путями. . .
Первый путь индукции апоптоза— внедрение в клетку-мишень перфорин-гранзимсодержащих гранул (рис. 3).
После специфического распознавания CD8+ Т-клеткой за счет своего антигенраспознающего рецептора чужеродного пептида, который находится на мембране клетки-мишени в составе молекулы гистосовместимости класса I, в CD8+ клетке (киллере) образуется специальный белок перфорин, способный формировать поры в мембране клетки-мишени. Кроме того, в CD8+ клетке образуется целое семейство сериновых протеаз, собранных в гранулы и названных гранзимами; наиболее важную роль из них играет гранзим В. На этапе летального удара ("поцелуя смерти") при взаимодействии Т-киллера и клетки-мишени перфорин повреждает ("пунктирует") мембрану клетки-мишени и растворимые гранзимы попадают внутрь клетки. Здесь они активируют целую серию цистеиновых протеаз — каспаз (сегодня их известно 10). В результате включается механизм апоптоза, который заканчивается активацией эндонуклеаз и фрагментацией ядра (см. рис. 3).
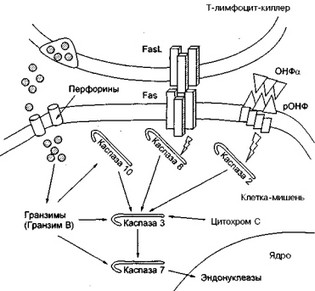
Рис. 3. Летальный удар ("поцелуй смерти") Т-лимфоцита-киллера
Гранзимы, в частности, гранзим В, активирует каспазу 10; взаимодействие Fas и FasL активирует каспазу 8; взаимодействие ОНФа и рецептора ОНФ активирует каспазу 2. Все они, в свою очередь, способны активировать каспазу 3. Эту каспазу активирует Также цитохром С. Каспаза 3 активирует каспазу 7, которая активирует ядерные эндонуклеазы, что заканчивается гибелью клетки. Кроме того, гранзим В способен напрямую активировать каспазу 3 и каспазу 7.
Таким образом, описанная выше дегрануляция цитотоксических Т-лимфоцитов и перфорининдуцированный апоптоз клеток являются конечным этапом специфического процесса, который требует предварительного взаимодействия Т-клеточного рецептора и комплекса (молекула ГКГ + пептид) для распознавания специфической клетки-мишени. До настоящего времени этот механизм разрушения клеток под влиянием Т-лимфоцитов-киллеров считается классическим при развитии реакции отторжения аллогенных трансплантатов, аутоиммунной патологии, разрушении вирусинфицированных и опухолевых клеток.
Второй путь индукции апоптоза —связывание Fas-лиганда, имеющегося на поверхности CD8+ клетки (киллера), с Fas-рецептором на поверхности соответствующей клетки-мишени.
Одновременно с первым, специфическим, механизмом киллинга, Т- киллер включает и второй, неспецифический, за счет связывания своего Fas-лиганда с Fas-рецептором клетки-мишени. Это также приводит к активации цйстеиновых протеаз и включению механизма апоптоза.
Подобный механизм гибели клетки развивается также под влиянием ЕК-клеток, фактора некроза опухолей-альфа и цитохрома С (последний высвобождается митохондриями стрессированной клетки- мишени) (см. рис. 3).
Источник: Г. Н. Дранник, «КЛИНИЧЕСКАЯ ИММУНОЛОГИЯ и АЛЛЕРГОЛОГИЯ» 1999
А так же в разделе «АПОПТОЗ »
- Глава 1 ОСНОВНЫЕ ЗАДАЧИ И ПРОБЛЕМЫ КЛИНИЧЕСКОЙ ИММУНОЛОГИИ
- Глава 2 ВРОЖДЕННЫЙ НЕСПЕЦИФИЧЕСКИЙ (ЕСТЕСТВЕННЫЙ) ИММУНИТЕТ
- Глава 3 СИСТЕМА КОМПЛЕМЕНТА
- КЛАССИЧЕСКИЙ ПУТЬ АКТИВАЦИИ СИСТЕМЫ КОМПЛЕМЕНТА
- АЛЬТЕРНАТИВНЫЙ ПУТЬ АКТИВАЦИИ СИСТЕМЫ КОМПЛЕМЕНТА
- БИОЛОГИЧЕСКИЕ ПОСЛЕДСТВИЯ АКТИВАЦИИ СИСТЕМЫ КОМПЛЕМЕНТА
- Глава 4 АНТИГЕНЫ
- Глава 5 ПРИОБРЕТЕННЫЙ СПЕЦИФИЧЕСКИЙ (АДАПТИВНЫЙ) ИММУНИТЕТ
- ОСНОВНЫЕ СВЕДЕНИЯ О СТРУКТУРЕ И ФУНКЦИИ СПЕЦИФИЧЕСКОГО (АДАПТИВНОГО) ИММУНИТЕТА
- СОЗРЕВАНИЕ Т- И В-ЛИМФОЦИТОВ
- ОТБОР (СЕЛЕКЦИЯ) ЛИМФОЦИТАРНОГО НАБОРА (РЕПЕРТУАРА); ОТБОР ЛИМФОЦИТОВ, НЕ РЕАГИРУЮЩИХ НА СОБСТВЕННЫЕ (SELF) АНТИГЕНЫ. СОЗДАНИЕ ТОЛЕРАНТНОСТИ
- Т-ЛИМФОЦИТЫ
- Т-ЛИМФОЦИТЫ-СУПРЕССОРЫ
- В-ЛИМФОЦИТЫ
- Т-НЕЗАВИСИМАЯ ПРОДУКЦИЯ АНТИТЕЛ
- Т-ЗАВИСИМАЯ ПРОДУКЦИЯ АНТИТЕЛ
- ДИФФЕРЕНЦИРОВКА Т-ЛИМФОЦИТОВ
- ДИФФЕРЕНЦИРОВКА В-ЛИМФОЦИТОВ
- Глава 6 ИММУНОГЛОБУЛИНЫ
- ИММУНОГЛОБУЛИН
- ИММУНОГЛОБУЛИН G
- ИММУНОГЛОБУЛИН А
- ИММУНОГЛОБУЛИН Е
- ИММУНОГЛОБУЛИН D
- Глава 7 ИММУННЫЕ КОМПЛЕКСЫ
- Глава 8 ЦИТОКИНЫ - РЕГУЛЯТОРЫ И ЭФФЕКТОРЫ ИММУННОЙ СИСТЕМЫ
- ИНТЕРЛЕЙКИНЫ
- РОСТОВЫЕ ФАКТОРЫ
- ОПУХОЛЬНЕКРОТИЗИРУЮЩИЕ ФАКТОРЫ
- ХЕМОКИНЫ
- КРАТКАЯ ХАРАКТЕРИСТИКА НАИБОЛЕЕ ВАЖНЫХ ХЕМОКИНОВ
- Глава 9 ИНТЕРФЕРОНЫ
- Глава 10 МОЛЕКУЛЫ КЛЕТОЧНОЙ АДГЕЗИИ (АДГЕЗИВНЫЕ МОЛЕКУЛЫ)
- Глава 11 ИММУННАЯ СИСТЕМА СЛИЗИСТЫХ ОБОЛОЧЕК
- ЛИМФОИДНАЯ ТКАНЬ, АССОЦИИРОВАННАЯ С ЖЕЛУДКОМ И КИШЕЧНИКОМ
- МИГРАЦИЯ ЛИМФОЦИТОВ ПРИ РАЗВИТИИ ИММУННОГО ОТВЕТА В ОБЛАСТИ СЛИЗИСТЫХ ОБОЛОЧЕК
- ТОЛЕРАНТНОСТЬ, ИНДУЦИРУЕМАЯ ВВЕДЕНИЕМ АНТИГЕНА per os
- СЕКРЕЦИЯ IgA
- ТРАНСПОРТ СЕКРЕТОРНОГО IgA
- ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ СЕКРЕТОРНОГО IgA
- ГАММА-, ДЕЛЬТА-Т-ЛИМФОЦИТЫ
- Глава 12 ГЛАВНЫЙ КОМПЛЕКС ГИСТОСОВМЕСТИМОСТИ: СТРУКТУРА И ФУНКЦИИ
- ВЗАИМОСВЯЗЬ АНТИГЕНОВ СИСТЕМЫ HLA С ПРЕДРАСПОЛОЖЕННОСТЬЮ К ЗАБОЛЕВАНИЯМ
- ОПРЕДЕЛЕНИЕ HLA-ФЕНОТИПА