Т-ЛИМФОЦИТЫ
Как упоминалось выше, часть иммунологически незрелых стволовых клеток мигрирует из костного мозга в вилочковую железу (тимус), где под влиянием тимического микроокружения, прежде всего эпителиальных клеток и гормонов тимуса, созревают до иммунокомпетентных Т-лимфоцитов. Созревание тимоцитов (незрелых лимфоцитов, попавших в тимус из костного мозга) происходит последовательно, по мере их перемещения из коркового слоя железы в мозговое вещество, т. е. ' из наружного слоя тимуса во внутренний.
У эмбриона человека тимус закладывается на 6-й неделе гестации и представляет собой эпителиальные клетки, окруженные мезенхимальной тканью. Лимфоидные клетки начинают определяться в тимусе человека на 13—14-й гестационной неделе. Тимус—источник Т-лим- фоцитов; подсчитано, что у крыс массой 100 г он продуцирует за 1 ч ~ 20 млн. лимфоцитов.
Созревает тимус к 12—15 годам, после чего начинается так называемая физиологическая инволюция: уменьшается зона коркового вещества, снижается тимоцитопоэз, продукция гормонов тимуса. Кроме возрастной, физиологической, необратимой инволюции различают еще акциденталъную, обратимую, инволюцию тимуса, которая может развиться в любом возрасте (особенно это важно в младшем детском возрасте) под влиянием различных стрессовых воздействий (психоэмоциональных, экологических, лекарственных и др.). Возрастная, также как и акцидентальная инволюция тимуса могут быть причиной развития вторичной иммунологической недостаточности.
Тимус продуцирует несколько гормонов, из них наиболее изучены следующие:
- Тимозин (фракция 5) содержит около 30 термостабильных полипептидов с различной молекулярной массой. По миграционным свойствам разделен на 3 класса: 1) альфа-тимозины — альфа-1, альфа-5 и альфа-7 (изоэлектр. ?очка —lt; 5,5); 2) бета-тимозины —:бета-3 и бета- 4 (5,5—7,0); 3) гамма-тимозины (gt; 7,0).
Таким образом, тимозин 5 включает: а) продукты разрушения ти- моцитов; б) лимфокины, продуцируемые тимоцитами; в) истинные гормоны, продуцируемые эпителиальными клетками тимуса — альфа-1, 5, 7- и бета-3, 4-полипептиды.
Тимозин 5 восстанавливает иммунный ответ, усиливает лимфоци- топоэз, вызывает лимфоцитоз, стимулирует антителообразование, противоопухолевый иммунитет, функцию Т-хелперов, супрессоров и киллеров. Тимозин, в отличие от других активных веществ тимуса, повышает уровень цГМФ и не влияет на цАМФ.
- Тимопоэт ин — I и II — различаются по двум аминокислотным остаткам. Активный центр тимопоэтина — пептид, соответствующий 20—41-й позициям аминокислот. Синтезирован минимальный фрагмент (32—36-я позиции аминокислот), сохраняющий активность тимопоэтина, — ТР5. Под влиянием тимопоэтина в претимических лимфоцитах повышается уровень цАМФ, а в периферических — цГМФ. На В-клетки гормон не действует. Тимопоэтин — иммуномодулятор, поскольку способен стимулировать и угнетать иммунные реакции. Видимо, для проявления биоактивности тимопоэтина необходимы еще какие-то вещества тимической природы, поскольку он не восстанавливает иммунокомпетентность неонатально тимэктомированных животных.
- Тимическийгуморальный фактор (ТГФ) — термолабильный полипептид, массой 3,2 • 103 D, имеет 31 аминокислотный остаток.
Стимулирует Т-систему: реакцию трансплантат против хозяина (РТПХ), реакцию бластной трансформации лимфоцитов (РБТЛ) на фитогемагглютинин (ФГА) и конканавалин (КонА), миграцию Т-кле- ток, отменяет вастинг-синдром у тимэктомированных животных, усиливает Т-киллеры и Т-хелперы. Клетка-мишень для него — Т-лимфо- цит. Механизм действия типичный для пептидных гормонов — через аденилатциклазную систему.
- Тимулин, или сывороточный тимический фактор (СТФ), — выделен из сыворотки крови; молекулярная масса— 8,57-102 D. Получен синтетический аналог. У человека постоянный уровень СТФ держится до 20 лет, затем начинает снижаться и к 50 годам исчезает. Нужен цинк для его синтеза. В сыворотке крови есть специальный белок-носитель для СТФ (похож на альбумин или преальбумин). Участвует в дифференцировке как претимических, так и посттимических лимфоцитов; in vitro стимулирует образование Т-супрессоров и Т-хелперов.
Итак, общим для всех перечисленных гормонов тимуса является их участие в процессах дифференцировки Т-лимфоцитов, возможно, на разных его этапах.
Группой украинских ученых во главе с И. А. Безвершенко из тимуса выделено так называемое лимфоцитозстимулирующее вещество (ЛСВ)— низкомолекулярный неполипептидный фактор. Состоит из 2 групп веществ с молекулярной массой 6—7- 102 D и 1,5—2-102 D. ЛСВ обнаружен в 5-й фракции, поскольку он не осаждается аммония сульфатом. Основная функция ЛВС — индукция пролиферации Т-лим- фоцитов как в тимусе, так и на периферии; активация репаративной регенерации Т-лимфоцитов (увеличение количества Т-клеток в тимусе) и повышение их миграции в селезенку и лимфатические узлы. Под влиянием ЛСВ образуются все классы Т-лимфоцитов — киллеры, супрессоры и хелперы. Эффект — дозозависимый. Малые дозы стимулируют, большие — угнетают. Ингибиция идет за счет увеличения числа Т-лимфоцитов-супрессоров. Это свойство ЛСВ — усиливать функцию Т-лимфоцитов-супрессоров — легло в основу механизма действия препарата "Вилозен", полученного в Украине из ЛСВ и используемого при лечении аллергических заболеваний для подавления продукции реагиновых (IgE) антител.
После окончания этапа антигеннезависимой дифференцировки в тимусе покоящиеся зрелые Т-лимфоциты, готовые к встрече с антигеном, через кровоток расселяются в определенных областях периферической лимфатической системы. Эти места получили название тимус- зависимых зон (Т-зон) вторичных лимфоидных органов. К ним относятся паракортикальная зона лимфатических узлов, богатая посткапиллярными венулами, обеспечивающими рециркуляцию лимфоцитов, а также периартериальные муфты лимфатических фолликулов селезенки (белая пульпа).
Как уже упоминалось, на поверхности всех Т-лимфоцитов имеется специальный инструмент, с помощью которого происходит распознавание чужеродного материала — Т-клеточный антигенраспознающий рецептор (ТАГРР) (рис. 1).
Этот рецептор представляет собой гетеродимер, состоящий из двух полипептидных цепей. Большая часть каждой из двух цепей рецептора находится вне клетки и свернута в виде двух доменов — вариабельного (V) и константного .(С). Большинство Т-лимфоцитов несет рецепторы, состоящие из альфа- и бета-цепей (рис. 1, а). Иногда такие Т-лим- фоциты называют "альфа-бета-Т-лимфоциты". Именно вариабельный
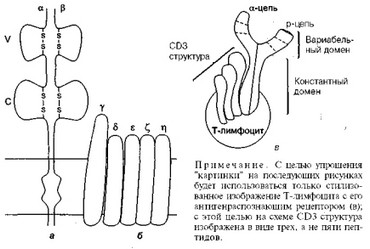
Рис. 1. Схематическое изображение строения Т-клеточного антиген- распознающего рецептора (ТАГГР) (объяснение в тексте)
домен обеспечивает существование огромного числа рецепторов разной специфичности, позволяющего распознать любой чужеродный материал, попавший в организм. Обязательным условием является наличие на одном Т-лимфоците рецептора (точнее, рецепторов), способного распознать только один антиген. Уже после распознавания для реализации полноценного иммунного ответа такой Т-лимфоцит подвергается пролиферации (делению), в результате чего из одного Т- лимфоцита образуется целый клон (группа) клеток, обладающих той же специфичность, что и первоначальный Т-лимфоцит.
У всех Т-лимфоцитов, несущих ТАГРР, последний нековалентно связан в единый комплекс с пятью трансмембранными белками: гамма (у), дельта (8), эпсилон (е), дзета (Q, эта (л) (см. рис. 1,6), которые в настоящее время обозначаются как единая дифференцировочная молекула CD3. Важнейшая функциональная роль молекулы CD3 состоит в том, что она участвует в передаче сигнала от собственно распознающих антиген альфа-, бета-цепей внутрь клетки, запуская процесс ее активации с последующей пролиферацией. Таким образом, ТАГРР представляет собой комплекс, состоящий из альфа-, бета-цепей и молекулы CD3, включающей пять мембранных белков, и весь этот комплекс следует рассматривать как единую функциональную структуру. (В со
ответствии с международной классификацией все основные антигенные маркеры лимфоцитов и других клеток иммунной системы сведены в группу и обозначены как кластеры дифференцировки, или CD (claster of differentiation). Набор различных CD на поверхности отдельной клетки составляет ее фенотип (поверхностную характеристику).
Суммируя выше изложенное, можно отметить следующее:
- На мембране зрелых покоящихся Т-лимфоцитов имеется ТАГРР с определенной антигенной специфичностью, не зависящей от того, встречался ли ранее организм с данным антигеном или нет;
- Встреча Т-лимфоцита со специфическим антигеном включает новый этап в жизни Т-лимфоцита— этап антигензависимой дифференцировки (в отличие от антигеннезависимой, которая прошла в тимусе);
- Распознавание специфического антигена приводит к активации Т-лимфоцита и последующей его пролиферации, заканчивающейся появлением в организме большого количества (клона) Т-лимфоцитов определенной специфичности, способной реализовывать' иммунный ответ.
Процесс распознавания чужеродного материала Т-лимфоцитом своеобразный, и отличается от процесса распознавания В-лимфоци- том. Для распознавания крупной чужеродной (бактериальная, вирусная и другие клетки) или аутологичной структуры Т-лимфоцитам необходим промежуточный этап, на котором макрофаг или другая ан- тиген-представляющая клетка специальным образом "подготавливает" чужеродный материал для распознавания. Этот процесс подготовки носит название процессинга (переваривания) и заключается в ферментативном расщеплении поглощенного макрофагом чужеродного материала.-Образующиеся в результате процессинга отдельные блоки, или пептиды, представляют собой определенной длины аминокислотные остатки — эпитопы чужеродного антигена. Именно эти пептиды и способны распознавать Т-лимфоциты в тот момент, когда они попадают на мембрану макрофага в сочетании с молекулами главного комплекса гистовместимости (ГКГ), или трансплантационными антигенами.
Более подробно механизмы распознавания будут описаны в специальной главе. .
Т-лимфоциты относятся, в основном, к долгоживущей и медленно рециркулирующей популяции лимфоидных клеток. Есть данные о том, что длительность жизни для некоторых Т-лимфоцитов составляет 15— 20 лет. Поскольку во взрослом состоянии замещаемость среди популяции Т-лимфоцитов невелика, то повреждения в этой части иммунной системы трудно восстанавливаются и имеют серьезные последствия. Например удаление тимуса в период онтогенеза приводит к
нарушению в иммунной системе, что проявляется нарушением клеточных реакций организма и снижением продукции иммуноглобулинов (антител) на тимусзависимые антигены. На долю Т-лимфоцитов в периферической крови приходится 55—75% от общего числа лимфоцитов, в селезенке — 60, а в лимфатическом узле — 70%.
Кроме ТАГРР, на поверхности всех Т-лимфоцитов также имеются:
- рецептор к эритроциту барана (в настоящее время его отождествляют с С02-структурой);
- рецептор к Т-митогенам: фитогемагглютинину (белок растительного происхождения, полученный из фасоли) и конканавалину А;
- рецепторы к интерлейкину 1, 2 (ИЛ-1, ИЛ-2);
- трансплантационные антигены класса I.
Это далеко не полный перечень антигенов и рецепторов, имеющихся на поверхности Т-лимфоцитов.
Многообразие функций, которые выполняют Т-лимфоциты в рамках иммунного ответа организма, связано с существованием различных субпопуляций Т-лимфоцитов, "запрограммированных" на реализацию конкретной задачи в целях поддержания иммуногомеостаза.
Т-клетки по их поверхностным гликопротеинам CD4 и CD8 подразделяют на две большие категории. Известно, что зрелые Т-клетки имеют только один из этих белков —либо CD4, либо CD8, но не оба.
В отличие от этого, незрелые Т-лимфоциты, находящиеся в тимусе,
¦ могут быть одновременно CD4- и СВ8-положительными (двупозитивными CD4+ CD8+).
CD4+ Лимфоциты получили название Т-лимфоциты-хелперы/индук-
торы, их количество в периферической крови достигает 55—60% от общего числа Т-лимфоцитов. CD8+ Лимфоциты названы Т-лимфоцитами- киллерами/супрессорами, их содержание в периферической крови составляет 20—30%. Обе субпопуляции Т-лимфоцитов относятся к так называемым иммунорегуляторным клеткам, и от их соотношения в организме во многом зависит сила иммунного ответа. По некоторым данным, соотношение иммунорегуляторных клеток генетически детерминировано и у каждого человека сугубо индивидуально. Все же, принято считать, что в среднем соотношение клеток в норме составляет 1,5—3,5. .
Молекулы CD4+ и CD8+ являются трансмембранными гликопротеинами, они относятся к так называемым адгезивным молекулам и участвуют в распознавании аутологичных молекул главного комплекса гистосовместимости, а также в механизмах передачи сигнала внутрь лимфоцита в момент кооперации Т-лимфоцитов и антигенпредстав- ляющих клеток при антигенном распознавании.
CD4+ Лимфоциты выполняют главным образом хелперную функцию, однако следует отметить, что при некоторых обстоятельствах CD4+ клетки могут выполнять и киллерную функцию. Выполняя свою основную хелперную функцию, они помогают, во-первых, В-клеткам превращаться в антителопродуцирующую плазматическую клетку; во-вторых, CD8+ лимфоцитам — в зрелую цитотоксическую Т-клетку; в-третьих, макрофагам осуществлять эффекты гиперчувствительности замедленного типа. Указанные функции Т-лимфоцитов-хелперов реализуются за счет того, что они в свою очередь разделяются на две субпопуляции — 1-го и 2-го типа, выполняющие разные хелперные функции за счет продукции разных цитокинов — интерлейкинов.
Т-Лимфоциты-хе/шерьг 1-го типа (Тх1) продуцируют гамма-интерферон (ИНФ), ИЛ-2 и альфа-опухольнекротизирующий фактор (ОНФ). Указанные цитокины активируют макрофаги, ЕК-клетки, созревание цитотоксических Т-лимфоцитов-киллеров, обеспечивая преимущественное развитие клеточного иммунного ответа, в том ¦ числе, при внутриклеточной инфекции.
Напротив, Т-лимфоциты-хелперы 2-го типа (Тх2) продуцируют ИЛ-4, ИЛ-5, ИЛ-10 и ИЛ-13, которые отвечают за развитие гуморального ответа, в том числе, за продукцию IgE. Кроме того, ИЛ-10 обладает ингибирующим эффектом по отношению к Txl.
Txl и Тх2 различаются не только по способности продуцировать различные цитокины, но и по наличию на своей поверхности различных активационных маркеров. Так, после активации на мембране Тх2 экспрессируется CD30 (молекула, относящаяся к семейству рецепторов для опухольнекротизирующего фактора), а на поверхности активированных Txl появляется LAG-3 (молекула, относящаяся к суперсемейству иммуноглобулинов).
Одним из важнейших регуляторных цитокинов, поддерживающих баланс между Т-хелперами 1-го и 2-го типа, является ИЛ-12, который продуцируется макрофагами. ИЛ-12 увеличивает количество Т-хел- перов 1-го типа, помогая таким образом хозяину защититься против микроорганизмов, которые контролируются клеточным иммунным ответом. Другой важный регуляторный компонент — это гамма-интерферон, который подавляет функционирование Т-хелперов 2-го типа.
Со своей стороны, Т-хелперы 2-го типа могут продуцировать ИЛ-10, который является супрессивным интерлейкином и подавляет функцию Т-лимфоцитов-хелперов 1-го типа. В настоящее время принято считать, что цитокины, которые продуцируются Txl и Тх2, используются как аутокринные факторы, и как факторы, способные вызыватьре- ципрокную супрессию (взаимное подавление функции).
На схеме 6 представлена дифференцировка Т-хелперов 1-го и 2-го
Дифференцировка Т-лимфоцитов-хелперов 1-го и 2-го типа (Тх1 и Тх2)
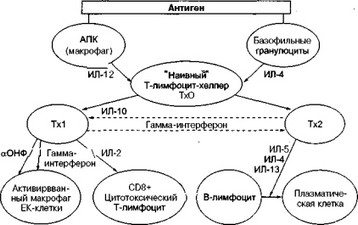
типа из покоящейся недифференцированной хелперной клетки. Как видно из схемы, на ранних этапах иммунного ответа под влиянием ИЛ-12, который продуцируется антигенпредставляющей клеткой (АПК), дифференцировка ТхО идет преимущественно в сторону созревания Txl, которые начинают продуцировать ИЛ-2, гамма-ИНФ и альфа-ОНФ. В случае воздействия на ТхО ИЛ-4, который продуцируется тканевыми базофилами (тучными клетками) и базофильными гранулоцитами крови, ТхО начинают дифференцироваться в Тх2 и продуцировать свой цитокиновый профиль: ИЛ-4, ИЛ-5, ИЛ-10, ИЛ-13. Гамма-ИНФ и ИЛ-10 способны рецикпрокно подавлять функционирование Txl и Тх2.
ИЛ-12 обладает способностью не только влиять на созревание Txl, но и стимулировать их пролиферацию как паракринный фактор. Точно так же действует и ИЛ-4 на Тх2: он сначала индуцирует дифферен- цировку Тх2, а затем уже как аутокринный фактор способствует их пролиферации.
Обнаружено, что Txl и Тх2 ответственны за развитие различных иммунопатологических реакций у человека. Так, например, функция Txl преобладает при развитии множественного (рассеянного) склероза, инсулинозависимого сахарного диабета, аутоиммунного тиреои- дита, болезни Крона, острого отторжения аллотрансплантата, при-
вычного невынашивания плода и др. В свою очередь, функция Тх2 преобладает при нормально протекающей беременности, трансплантационной толерантности, идиопатическом легочном фиброзе, прогрессирующем системном склерозе, у ВИЧ-инфицированных больных с быстрым прогрессированием заболевания, а также при аллергической патологии.
Таким образом, Т-лимфоциты-хелперы 1-го и 2-го типа представляют собой важнейшие субпопуляции Т-лимфоцитов, от функционального баланса которых зависит направленность иммунного ответа в норме и особенности клинических проявлений при развитии иммунопатологии. На этом основании Т-хелперы получили название "дирижеры иммунного ответа". В табл. 2 приведена сравнительная характеристика Т-лимфоцитов-хелперов обоих типов.
Таблица 2. Сравнительная характеристика Т-лимфоцитов-хелперов 1-го и 2-го типа
|
Свойства |
Тх1 |
Тх2 |
|
Продукция ИЛ-2, ИНФ, альфа-ОНФ |
+ |
- |
|
Продукция ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10 и ИЛ-13 |
- |
+ |
|
Усиление клеточного иммунитета и гиперчувствительности замедленного типа |
+ |
- |
|
Усиление продукции антител |
- |
+ |
|
Стимуляция под влиянием ИЛ-12 - |
+ |
- |
|
Стимуляция под влиянием ИЛ-4 |
- |
+ |
Другая субпопуляция Т-лимфоцитов, меньшая по количеству, несет на своей поверхности отличительный признак в виде молекулы CD8 и относится, как уже упоминалось, кТ-лимфоцитам-киллерам/су- прессорам. Такое двойное название означает, что эта субпопуляция ' Т-лимфоцитов может дифференцироваться либо в Т-киллер (цитотоксический Т-лимфоцит), либо в Т-супрессор и выполнять различные функции в зависимости от потребностей организма.
До недавнего времени безоговорочно признавалось существование CD8+ супрессорных клеток, которые вместе с CD4+ клетками относились к иммунорегуляторным субпопуляциям. Однако, данные последних лет, особенно открытие Т-хелперов 1-го и 2-го типа, внесли некоторые сомнения в существование постулированных ранее супрессорных CD8+ клеток, хотя функциональных доказательств предостаточно. В связи с этим предполагается, что способность Т-лимфоцитов-хелпе- ров 2-го типа продуцировать супрессорный ИЛ-10 и является тем моментом, который позволяет Т-лимфоцитам-хелперам 1-го и 2-го типа без участия других клеток реализовывать регуляторный потенциал иммунного ответа.
Будущие исследования покажут, существует ли морфологически очерченный тип Т-лимфоцитов-супрессоров, или иммунорегуляторная роль будет полностью закреплена за соотношением субпопуляций Т- лимфоцитов-хелперов 1-го и 2-го типа. Тем не менее, сегодня продолжают считать, что GD8+ клетки киллеры/супрессоры существуют, а соотношение Тх:Тс является важным иммунорегуляторным индексом, играющим существенную роль в поддержании нормального иммунного ответа. Более того, недавно получены доказательства о существовании морфологической структуры на поверхности CD8+ клеток, с помощью которой эту субпопуляцию можно разделить на две, функционально отличающиеся между собой группы клеток: киллеры и супрессоры. По имеющимся данным, для Т-киллеров характерен поверхностный фенотип CD8+CD28+, а для Т-супрессоров — CD8+CD28- (М. Е. North и соавторы, 1998).
Итак, прежде всего, охарактеризуем CD8+ лимфоциты, выполняющие цитотоксические функции. Эти лимфоциты реализуют специфические клеточные реакции иммунитета: участвуют в механизмах отторжения аллотрансплантатов, реакциях аутоиммунитета, разрушают вирусинфицированные и опухолевые клетки.
Таким образом, Т-лимфоцит-киллер — основная эффекторная клетка клеточно-опосредованного иммунитета, которая осуществляет лизис мишеней, обеспечивает генетическое постоянство внутренней среды организма. -
Напомним, что в периферической крови и во вторичных лимфоидных органах Т-киллер находится в состоянии покоя, — так называемая зрелая покоящаяся CD8+ клетка. Для того, чтобы произошла ее дифференцировка в зрелый Т-киллер, способный осуществлять кил- линговый эффект, требуется несколько условий. Во-первых, нужно распознать чужеродный антиген; во-вторых, требуется время для создания клона специфических Т-киллеров, способных оказать ощутимый эффект. Для распознавания чужеродного антигена у CD8+ клетки, так же, как и у CD4+, есть Т-клеточный антигенраспознающий рецептор в комплексе с СОЗ-структурой. Точно так же, как и в случае с CD4+ клеткой (хелпером), CD8+ клетка (киллер) распознает не весь чужеродный антиген, а его блоки, так называемые доминантные пептиды, которые находятся на поверхности антигенпредставляющей клетки в сочетании с молекулами ГКГ. Однако, существует принципиальное различие в "работе" CD4+ клеток (хелперов) и CD8+ клеток (киллеров) при распознавании антигенов.
Так, CD4+ клетки (Т-лимфоциты-хелперы) могут распознать чужеродный пептид только в том случае, если он находится в сочетании (презентируется) с молекулами гистосовместимости' класса II на поверхности антигенпрезентирующей клетки. В норме в организме таких клеток немного — это моноциты-макрофаги, В-лимфоциты и дендритные клетки, обладающие способностью поглощать попавший в организм чужеродный материал, перерабатывать (процессировать) его с помощью целого ряда ферментов, разрезая антиген на блоки — пептиды, а затем транспортировать их из глубины клетки на ее поверхность в сочетании с молекулами гистосовместимости класса II. Только после этого CD4+ клетка (хелпер) сможет распознать эти чужеродные, так называемые экзогенные, пептиды; это повлечет за собой активацию и пролиферацию CD4+ клеток с последующей их дифференцировкой на Т-хелперы 1-го и 2-го типа, о чем уже упоминалось.
Совсем иначе осуществляется распознавание пептидов CD8+ клеткой (Т-лимфоцитом-киллером). Основное отличие состоит в том, что пептид подается для распознавания (презентируется) в составе молекулы гистосовместимости класса I, а не класса II, как для хелперов. Это очень важный момент, поскольку молекулы гистосовместимости класса I присутствуют на всех ядерных клетках организма. Исходя из этого, все изменения гомеостаза организма, происходящие на внутриклеточном уровне, будут отражаться на мембране клетки в виде так называемых эндогенных пептидов, находящихся в составе молекул гистосовместимости класса I. То или иное изменение гомеостаза клетки превращает ее в чужеродную (например мутация, поражение вирусом и др.), CD8+ клетка (киллер) распознает это по пептидам, проактивируется и разрушит такую измененную клетку. Таким образом, CD8+ Т-клетка (киллер), распознающая эндогенные пептиды в составе молекул гистосовместимости класса I, которые имеются на мембране всех ядерных клеток организма, выполняет своеобразную цензорную функцию, позволяющую иммунной системе осуществлять контроль за постоянством внутренней среды организма. Следует добавить, что с помощью молекул гистосовместимости класса I презентируются также экзогенные пептиды, "сделанные" из внутриклеточных паразитов, например, вирусов.
Существует еще одно условие, необходимое для созревания цитотоксических CD8+ клеток: после распознавания чужеродного пептида эти клетки должны получить дополнительный сигнал от CD4+ клеток (хелперов), который позволит им делиться (пролиферировать), в результате чего из одной клетки образуется целый клон (группа) клеток, обладающих одной специфичностью и достаточным потенциалом для реализации клеточного иммунного ответа. Отсюда ясно, насколько важна способность клеток пролиферировать. Сигнал к пролиферации CD4+ клетка (хелпер) подает с помощью ИЛ-2, который она продуцирует; для восприятия этого сигнала на CD8+ клетке (киллере) есть рецептор к ИЛ-2. Более точная хронология событий состоит в следующем.
После поглощения антигена, активированный макрофаг среди прочего продуцирует ИЛ-1, одна из основных задач которого заключается в том, чтобы "заставить" Т-лимфоциты-хелперы продуцировать ИЛ-2. Одновременно под влиянием ИЛ-1 на поверхности лимфоцитов появляется рецептор к ИЛ-2. В том случае, если иммунный ответ пойдет по клеточному пути, то Т-лимфоцит-киллер после распознавания чужеродного пептида получит дополнительный сигнал в виде ИЛ-2 и начнет пролиферировать.
Рассмотрим цитотоксическую реакцию на примере разрушения ви- русинфицированных клеток (рис. 2). При появлении в организме ви-
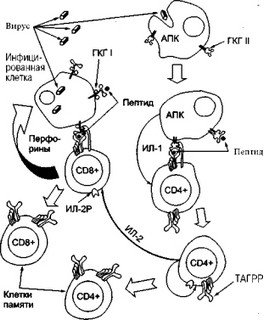
Рис. 2. Этапы "созревания” цитотоксических Т-лимфоцитов (CD8+ клетки) . при вирусной инфекции (объяснение в тексте).
русифицированных клеток, CD8+ Т-лимфоциты (киллеры) распознают вирусный антиген, который представляется им совместно с молекулой ГКГ класса I на поверхности этой клетки. В свою очередь CD4+ Т-лимфоцит (хелпер) распознает вирусный антиген, который представляется ему с молекулой ГКГ класса II антигенпредставляющей клетки (АПК), например макрофага. Одновременно макрофаг продуцирует ИЛ-1, что запускает интерлейкиновый каскад. Хелперный Т-лимфо- цит секретирует ИЛ-2, который позволяет предшественнику CD8+ цитотоксической Т-клетки пролиферировать, что приводит к образовы- ванию клона вирусспецифических клеток-киллеров. В последующем эти цитотоксические Т-клетки разрушают вирусинфицированные клетки. Одновременно с клеточным ответом на вирус развивается и гуморальный ответ, заканчивающийся продукцией специфических, антител. Здесь также очень важна роль CD4+ клеток (хелперов). Более подробно об этом будет сказано ниже.
Механизм цитолитического действия Т-лимфоцитов-киллеров в настоящее время представляется следующим образом: на первом этап е (программирования лизиса) между клеткой-эффектором (киллером) и клеткой-мишенью устанавливается специфический контакт; на втором этапе (летального удара) клетки-киллеры оказывают литическое действие на клетки-мишени; на третьем (заключительном) этапе осуществляется непосредственное повреждение клеток-мишеней. В цитотоксической реакции разрушаются только клетки-мишени: кил- лерные клетки после летального удара отделяются от клеток-мишеней. Таким образом, Т-киллеры только запускают цитолитическую реакцию, но не участвуют в ' непосредственном разрушении клеток-мишеней. Сама киллерная клетка может участвовать в последовательном разрушении нескольких клеток-мишеней, оставаясь при этом неповрежденной и функционально активной.
Убитые вирусные вакцины не активируют CD8+ Т-клетки (киллеры) потому, что в этом случае вирус не реплицируется в пораженной клетке, и, следовательно, вирусные эпитопы (пептиды) не презентиру- ются в ассоциации с молекулами ГКГ класса I и не распознаются CD8+ клетками.
Следует учитывать, что активирование Т-клетки не является простой функцией "включения-выключения". Связывание эпитопов (пептидов) Т-клеточным распознающим рецептором может приводить либо к полной активации Т-клеток, либо к частичной, либо, наконец, не вызывать активацию. Все это зависит от того, на каком этапе прерываются сигналы трансдукции, идущие внутрь клетки, т. е. насколько данный пептид-эпитоп может индуцировать этот , сигнал трасдукции.
Для большей стабилизации взаимодействующих между собой Т- лимфоцитов и антигенпрезентирующих клеток в иммунном ответе необходимы также так называемые ко-стимулирующиесигналы. Они состоят в следующем: на Т-лимфоцитах имеется белок LFA-1, который связывается с соответствующим белком ICAM-1 на антигенпрезенти- рующей клетке. Кроме того, на поверхности Т-лимфоцитов есть белок CD28, который соединяется с белком CD80 на антигенпрезентирующих клетках. Для полной активации Т-лимфоцитов связь CD28 и CD80 крайне необходима. Если этого контакта не будет, то наступит анергия или апоптоз Т-лимфоцита.
Источник: Г. Н. Дранник, «КЛИНИЧЕСКАЯ ИММУНОЛОГИЯ и АЛЛЕРГОЛОГИЯ» 1999
А так же в разделе «Т-ЛИМФОЦИТЫ »
- Глава 1 ОСНОВНЫЕ ЗАДАЧИ И ПРОБЛЕМЫ КЛИНИЧЕСКОЙ ИММУНОЛОГИИ
- Глава 2 ВРОЖДЕННЫЙ НЕСПЕЦИФИЧЕСКИЙ (ЕСТЕСТВЕННЫЙ) ИММУНИТЕТ
- Глава 3 СИСТЕМА КОМПЛЕМЕНТА
- КЛАССИЧЕСКИЙ ПУТЬ АКТИВАЦИИ СИСТЕМЫ КОМПЛЕМЕНТА
- АЛЬТЕРНАТИВНЫЙ ПУТЬ АКТИВАЦИИ СИСТЕМЫ КОМПЛЕМЕНТА
- БИОЛОГИЧЕСКИЕ ПОСЛЕДСТВИЯ АКТИВАЦИИ СИСТЕМЫ КОМПЛЕМЕНТА
- Глава 4 АНТИГЕНЫ
- Глава 5 ПРИОБРЕТЕННЫЙ СПЕЦИФИЧЕСКИЙ (АДАПТИВНЫЙ) ИММУНИТЕТ
- ОСНОВНЫЕ СВЕДЕНИЯ О СТРУКТУРЕ И ФУНКЦИИ СПЕЦИФИЧЕСКОГО (АДАПТИВНОГО) ИММУНИТЕТА
- СОЗРЕВАНИЕ Т- И В-ЛИМФОЦИТОВ
- ОТБОР (СЕЛЕКЦИЯ) ЛИМФОЦИТАРНОГО НАБОРА (РЕПЕРТУАРА); ОТБОР ЛИМФОЦИТОВ, НЕ РЕАГИРУЮЩИХ НА СОБСТВЕННЫЕ (SELF) АНТИГЕНЫ. СОЗДАНИЕ ТОЛЕРАНТНОСТИ
- АПОПТОЗ
- Т-ЛИМФОЦИТЫ-СУПРЕССОРЫ
- В-ЛИМФОЦИТЫ
- Т-НЕЗАВИСИМАЯ ПРОДУКЦИЯ АНТИТЕЛ
- Т-ЗАВИСИМАЯ ПРОДУКЦИЯ АНТИТЕЛ
- ДИФФЕРЕНЦИРОВКА Т-ЛИМФОЦИТОВ
- ДИФФЕРЕНЦИРОВКА В-ЛИМФОЦИТОВ
- Глава 6 ИММУНОГЛОБУЛИНЫ
- ИММУНОГЛОБУЛИН
- ИММУНОГЛОБУЛИН G
- ИММУНОГЛОБУЛИН А
- ИММУНОГЛОБУЛИН Е
- ИММУНОГЛОБУЛИН D
- Глава 7 ИММУННЫЕ КОМПЛЕКСЫ
- Глава 8 ЦИТОКИНЫ - РЕГУЛЯТОРЫ И ЭФФЕКТОРЫ ИММУННОЙ СИСТЕМЫ
- ИНТЕРЛЕЙКИНЫ
- РОСТОВЫЕ ФАКТОРЫ
- ОПУХОЛЬНЕКРОТИЗИРУЮЩИЕ ФАКТОРЫ
- ХЕМОКИНЫ
- КРАТКАЯ ХАРАКТЕРИСТИКА НАИБОЛЕЕ ВАЖНЫХ ХЕМОКИНОВ
- Глава 9 ИНТЕРФЕРОНЫ
- Глава 10 МОЛЕКУЛЫ КЛЕТОЧНОЙ АДГЕЗИИ (АДГЕЗИВНЫЕ МОЛЕКУЛЫ)
- Глава 11 ИММУННАЯ СИСТЕМА СЛИЗИСТЫХ ОБОЛОЧЕК
- ЛИМФОИДНАЯ ТКАНЬ, АССОЦИИРОВАННАЯ С ЖЕЛУДКОМ И КИШЕЧНИКОМ
- МИГРАЦИЯ ЛИМФОЦИТОВ ПРИ РАЗВИТИИ ИММУННОГО ОТВЕТА В ОБЛАСТИ СЛИЗИСТЫХ ОБОЛОЧЕК
- ТОЛЕРАНТНОСТЬ, ИНДУЦИРУЕМАЯ ВВЕДЕНИЕМ АНТИГЕНА per os
- СЕКРЕЦИЯ IgA
- ТРАНСПОРТ СЕКРЕТОРНОГО IgA
- ФУНКЦИОНАЛЬНОЕ ЗНАЧЕНИЕ СЕКРЕТОРНОГО IgA
- ГАММА-, ДЕЛЬТА-Т-ЛИМФОЦИТЫ
- Глава 12 ГЛАВНЫЙ КОМПЛЕКС ГИСТОСОВМЕСТИМОСТИ: СТРУКТУРА И ФУНКЦИИ
- ВЗАИМОСВЯЗЬ АНТИГЕНОВ СИСТЕМЫ HLA С ПРЕДРАСПОЛОЖЕННОСТЬЮ К ЗАБОЛЕВАНИЯМ
- ОПРЕДЕЛЕНИЕ HLA-ФЕНОТИПА