Проводящие пути головного и спинного мозга
Импульсы, возникающие при воздействии на рецепторы, передаются по отросткам нейронов к их телам. Благодаря многочисленным синапсам нейроны контактируют между собой, образуя цепи, по которым нервные импульсы распространяются только в определенном направлении — от чувствительных нейронов через вставочные к эффекторным. Это обусловлено морфофункциональными особенностями синапсов, которые проводят возбуждение только в одном направлении — от пресинаптической мембраны к постсинаптической.
По одним цепям нейронов импульс распространяется центростремительно от места его возникновения в коже, слизистых оболочках, органах движения, стенках сосудов к спинному или головному мозгу. По другим цепям нейронов импульс проводится центробежно из мозга на периферию, к рабочим органам — мышцам, железам. В процессе эволюции и прогрессирующего развития центральной нервной системы строение рефлекторных дуг усложнилось. Возникли сложные рефлекторные дуги, образованные нейронами, расположенными в вышележащих сегментах спинного мозга, в базальных ядрах головного мозга и в коре полушарий большого мозга. Отростки нейронов направляются из спинного мозга вверх к различным структурам головного мозга, образуя пучки,
соединяющие между собой нервные центры. Эти пучки составляют проводящие пути. Проводящие пути — это совокупность тесно расположенных нервных волокон, соединяющих различные центры головного и спинного мозга, проходящих в определенных зонах их белого вещества и проводящих определенные нервные импульсы.
В спинном и головном мозге выделяют три группы проводящих путей (нервных волокон): ассоциативные, комиссуральные и проекционные.
Ассоциативные нервные волокна (короткие и длинные) соединяют между собой группы нейронов (нервные центры), расположенные в одной половине мозга. Короткие (внутридолевые) нервные волокна соединяют близлежащие участки серого вещества и располагаются, как правило, в пределах одной доли мозга. Длинные (междолевые) ассоциативные пучки соединяют между собой участки серого вещества, расположенные на значительном расстоянии один от другого, обычно в различных долях. К ним относятся: верхний продольный пучок, соединяющий кору лобной доли с теменной и затылочной; нижний продольный пучок, связывающий серое вещество височной доли с затылочной; крючковидный пучок, соединяющий кору в области лобного полюса с передней частью височной доли.
В спинном мозге ассоциативные волокна соединяют между собой нейроны, расположенные в различных сегментах. Эти волокна образуют собственные пучки спинного мозга (межсегментар- ные пучки), которые располагаются вблизи серого вещества.
Комиссуральные (спаечные) нервные волокна соединяют аналогичные центры (серое вещество) правого и левого полушарий большого мозга, образуя мозолистое тело, спайку свода и переднюю (белую) спайку. Мозолистое тело соединяет между собой новые, более молодые отделы коры большого мозга правого и левого полушарий. В каждом полушарии волокна расходятся веерообразно, образуя лучистость мозолистого тела.
В передней спайке проходят волокна, которые соединяют между собой участки коры височных долей обоих полушарий и принадлежат обонятельному мозгу (серое вещество гиппокампов и височных долей обоих полушарий).
Проекционные нервные волокна (проводящие пути) соединяют спинной мозг с головным, ядра мозгового ствола с базальными ядрами и корой полушарий большого мозга (восходящие пути), а также головной мозг со спинным (нисходящие пути).
Восходящие проекционные пути, афферентные, чувствительные. По ним к коре большого мозга поступают нервные импульсы, возникшие в результате воздействия на организм различных факторов внешней среды, включая импульсы, идущие от органов
чувств, опорно-двигательного аппарата, внутренних органов и сосудов. В зависимости от этого восходящие проекционные пути делятся на три группы: экстероцептивные, проприоцептивные, интероцептивные.
- Экстероцептивные проводящие пути несут импульсы от кожного покрова (болевые, температурные, осязания и давления), от органов чувств (зрительные, слуховые, вкусовые, обонятельные). Проводящий путь болевой и температурной чувствительности (латеральный спиноталамический путь) состоит из трех нейронов. Рецепторы первого (чувствительного) нейрона, воспринимающие указанные раздражения, располагаются в коже и слизистых оболочках. Тело первого нейрона лежит в спинномозговом узле. Центральный отросток в составе заднего корешка направляется в задний рог спинного мозга и заканчивается синапсами на клетках второго нейрона. Аксоны вторых нейронов, тела которых лежат в заднем роге, через переднюю серую спайку переходят на противоположную сторону спинного мозга, входят в боковой канатик, образуя латеральный спиноталамический путь. Он поднимается в продолговатый мозг, проходит в покрышке моста, покрышке среднего мозга и заканчивается в таламусе. Аксоны клеток таламуса (третий нейрон) направляются к внутренней зернистой пластинке (IV слой коры) постцентральной извилины, где находится корковый конец анализатора общей чувствительности.
Проводящий путь осязания и давления (передний спиноталамический путь) несет импульсы от рецепторов кожи к клеткам коры постцентральной извилины. Ход волокон первого нейрона этого пути аналогичен такому же нейрону латерального спиноталамического пути. Большинство аксонов второго нейрона переднего спиноталамического пути также переходят на противоположную сторону спинного мозга через переднюю серую спайку, входят в передний канатик и в его составе следуют вверх к таламусу. Часть волокон второго нейрона идет в составе заднего канатика спинного мозга вместе с аксонами проводящего пути проприоцептмв- ной чувствительности коркового направления.
- Проприоцептивные пути проводят импульсы от мышц, сухожилий, суставных капсул, связок. Эти пути несут информацию о положении частей тела, объеме движений, мышечном тонусе, степени натяжения сухожилий. Проводящий путь проприоцептивной чувствительности коркового направления несет импульсы мышеч- но-суставного чувства к коре постцентральной извилины. Проприо- цептивная чувствительность позволяет человеку оценивать положение частей своего тела в пространстве, анализировать собственные сложные движения и дает возможность проводить целенаправленную их коррекцию. Тела первого нейрона этого пути также лежат в
спинномозговом узле. Аксоны этих нейронов в составе заднего корешка, не входя в задний рог, направляются в задний канатик, где образуют тонкий и клиновидный пучки. Эти пучки следуют вверх в продолговатый мозг к тонкому и клиновидному ядрам. Аксоны вторых нейронов, выходящие из этих ядер, переходят на противоположную сторону, образуя медиальную петлю. Затем они проходят через покрышку моста и покрышку среднего мозга и заканчиваются в таламусе синапсами на телах третьих нейронов. Аксоны последних направляются в кору постцентральной извилины, где заканчиваются синапсами на нейронах IV слоя коры. Другая часть волокон вторых нейронов по выходе из тонкого и клиновидного ядер направляется через нижнюю мозжечковую ножку в кору червя на своей стороне мозжечка. Третья часть волокон переходит на противоположную сторону и также через нижнюю мозжечковую ножку направляется к коре червя на противоположной стороне мозжечка. Они несут проприоцептивные импульсы к мозжечку для коррекции подсознательных движений опорно-двигательного аппарата. Кроме описанного, имеются проприоцептивные передний и задний спиномозжечковые пути, которые несут в мозжечок информацию о состоянии опорно-двигательного аппарата и двигательных центров спинного мозга. - Интероцептивные пути проводят импульсы от внутренних органов и сосудов. Расположенные в них рецепторы (механо-, баро-, хемо-) воспринимают информацию о состоянии гомеостаза (интенсивности обменных процессов, химическом составе крови, тканевой жидкости, давлении в сосудах и т.д.).
Нисходящие проводящие пути проводят импульсы от коры полушарий большого мозга и подкорковых центров к ядрам мозгового ствола и двигательным ядрам передних рогов спинного мозга. Эти пути разделяются на две группы: пирамидные и экстрапира- мидные. Первые являются главными двигательными путями. Они несут через соответствующие двигательные ядра головного и спинного мозга импульсы из коры полушарий большого мозга к скелетным мышцам головы, шеи, туловища, конечностей. Экстра- пирамидные пути несут импульсы от подкорковых центров и различных отделов коры к двигательным ядрам черепных и спинномозговых нервов, затем к мышцам, а также другим нервным центрам ствола головного мозга и спинному мозгу.
Главный двигательный, или пирамидный, путь представляет собой систему нервных волокон, по которым произвольные двигательные импульсы от гигантопирамидальных невроцитов (пирамидальных клеток Беца), расположенных в коре предцентральной извилины (V слой), направляются к двигательным ядрам черепных нервов и к передним рогам спинного мозга, а от них к ске
летным мышцам. В зависимости от направления и расположения волокон пирамидный путь делится на три части: корково-ядерный путь, идущий к ядрам черепных нервов; латеральный и передний корковоспинномозговые (пирамидные) пути, идущие к ядрам передних рогов спинного мозга (рис. 105).
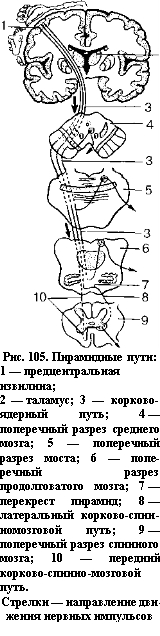 Корково-ядерный путь представляет собой пучок аксонов гигантопирамидальных клеток предцент- ральной извилины. Этот путь проходит через колено внутренней капсулы, основание ножки мозга. Волокна корково-ядерного пути переходят на противоположную сторону к двигательным ядрам черепных нервов, где заканчиваются синапсами на их нейронах. Аксоны двигательных нейронов указанных ядер выходят из мозга в составе соответствующих черепных нервов и направляются к скелетным мышцам головы и шеи.
Корково-ядерный путь представляет собой пучок аксонов гигантопирамидальных клеток предцент- ральной извилины. Этот путь проходит через колено внутренней капсулы, основание ножки мозга. Волокна корково-ядерного пути переходят на противоположную сторону к двигательным ядрам черепных нервов, где заканчиваются синапсами на их нейронах. Аксоны двигательных нейронов указанных ядер выходят из мозга в составе соответствующих черепных нервов и направляются к скелетным мышцам головы и шеи.
Латеральный и передний корковоспинномозговые (пирамидные) пути начинаются от гигантопирамидальных невроцитов предцентральной извилины. Волокна этого пути направляются к внутренней капсуле, проходят через переднюю часть ее задней ножки, затем через основание ножки мозга и моста, переходят в продолговатый мозг, образуя его пирамиды. На границе продолговатого и спинного мозга часть волокон корково-спинномозгового пути переходит на противоположную сторону, продолжается в
боковой канатик спинного мозга
(латеральный корково-спинномозговой путь) и постепенно заканчивается в передних рогах спинного мозга синапсами на их двигательных клетках. Волокна корково-спинномозгового пути, не
переходящие на противоположную сторону на границе продолговатого мозга со спинным, спускаются вниз в составе переднего канатика спинного мозга, образуя передний корково-спинномозговой путь. Эти волокна посегментно переходят на противоположную сторону через белую спайку спинного мозга и заканчиваются синапсами на двигательных невроцитах передних рогов противоположной стороны спинного мозга. Аксоны клеток передних рогов выходят из спинного мозга в составе переднего корешка и иннервируют скелетные мышцы. Итак, все пирамидные пути являются перекрещенными.
Экстрапирамидные проводящие пути являются филогенетически более старыми, чем пирамидные. Они имеют множество связей с клетками и ядрами ствола мозга и с корой большого мозга, которая контролирует и управляет экстрапирамидной системой. В связи с этим общим началом экстрапирамидных путей можно считать кору полушарий большого мозга, а местам, где они оканчиваются, — ядра мозгового ствола и передних рогов спинного мозга. Влияние коры полушарий большого мозга осуществляется через ряд образований: мозжечок, красные ядра, ретикулярную форма- .» цию, связанную с таламусом и полосатым телом через вестибу- лярные ядра. Одной из функций красного ядра является поддер- f жание мышечного тонуса, необходимого для непроизвольного ‘ удержания тела в равновесии. От красного ядра нервные импуль- | сы направляются в двигательные ядра передних рогов спинного мозга (красноядерно-спинномозговой путь). !
В осуществлении координации движений человека при нарушении равновесия важную роль играет преддверно-спинномозго- вой путь, который соединяет вестибулярные ядра с передними рогами спинного мозга. Первый нейрон этого пути залегает в ядрах VIII пары черепных нервов. Эти ядра соединены с мозжечком и посредством заднего продольного пучка — с двигательны- I ми ядрами III, IV, VI пар черепных нервов. Это обеспечивает | сохранение положения глазного яблока при движениях головы и ; шеи. Аксоны вторых нейронов преддверно-спинномозгового пути ; спускаются вниз в составе переднего канатика спинного мозга и ' заканчиваются синапсами на двигательных клетках передних ро- гов спинного мозга. Нейроны ретикулярной формации обеспе- •: чиваюг связь преддверно-спинномозгового пути с базальными ! ядрами.
Кора полушарий большого мозга осуществляет управление функциями мозжечка, участвующего в координации движений, 1 через мост по корково-мостомозжечковому пути. я
Таким образом, проводящие пути головного и спинного моз- I га устанавливают связи между афферентными и эфферентными I
(эффекторными) центрами, замыкают сложные рефлекторные дуги в теле человека. Одни проводящие пути несут нервные импульсы к ядрам филогенетически более старых отделов мозга, лежащих в мозговом стволе и обеспечивающих функции, которые обладают определенным автоматизмом, без участия сознания, хотя и под контролем полушарий большого мозга. Другие проводящие пути замыкаются с участием функций коры полушарий большого мозга (высших отделов центральной нервной системы) и обеспечивают произвольные действия органов и систем органов. Проводящие пути функционально объединяют организм в единое целое, обеспечивают согласованность его действий.
Возрастные особенности структуры и функций органов нервной системы
После рождения ребенка в первую очередь происходит миелинизация спинномозговых нервов, затем проводящих путей спинного мозга и ствола головного мозга. Основная масса волокон черепных нервов миелинизируется к 1,5—2 годам. В среднем к 3 годам основная масса нервных волокон миелинизи- рована, остальные завершают этот процесс к 6 годам. Относительно поздно завершают процесс миелинизации тангенциальные волокна коры полушарий большого мозга (к 30—40 годам). В процессе миелинизации происходит концентрация ионных каналов в области перехватов Ранвье. Повышается возбудимость и лабильность нервных волокон. Так, у новорожденных нерв способен проводить только 4—10 имп/с, в то время как у взрослых — 300—1000 имп/с.
Тормозные механизмы центральной нервной системы формируются в онтогенезе за счет развития тормозных нейронов. Становление тормозных механизмов существенно повышает способность к концентрации возбуждения, ограничивая его иррадиацию. С появлением тормозных механизмов безусловные рефлексы становятся более точными и локализованными.
Спинной мозг. В течение первых трех месяцев внутриутробной жизни спинной мозг занимает позвоночный канал на всю его длину. В дальнейшем позвоночник растет быстрее, чем спинной мозг. Поэтому нижний конец спинного мозга поднимается («восходит») в позвоночном канале. У новорожденного ребенка нижний конец спинного мозга находится на уровне III поясничного позвонка, у взрослого человека — на уровне — II поясничного позвонка. В связи с «восхождением» (относительным укорочением) спин
ного мозга в позвоночном канале корешки спинномозговых нервов удлиняются, принимают косое, а в нижних отделах — вертикальное положение. Корешки спинномозговых нервов, идущие к крестцовым отверстиям, образуют вокруг конечной нити пучок, получивший название «конский хвост».
Спинной мозг новорожденного имеет длину 14 см. К 2 годам длина спинного мозга достигает 20 см, а к 10 годам, по сравнению с периодом новорожденное™, удваивается. Быстрее всего растут грудные сегменты спинного мозга. Масса спинного мозга у новорожденного составляет около 5,5 г, у детей 1 -го года — около 10 г. КЗ годам масса спинного мозга превышает 13 г, к 7 годам равна примерно 19 г. У новорожденного центральный канал шире, чем у взрослого. Уменьшение его просвета происходит главным образом в течение 1—2 годов, а также В/более поздние возрастные периоды, когда наблюдается увеличение массы серого и белого вещества. Объем белого вещества спинного мозга возрастает быстро, особенно за счет собственных пучков сегментарного аппарата, формирование которого происходит в более ранние сроки по сравнению со сроками формирования проводящих путей, образующих надсегментарный аппарат мозга.
Вопросы для повторения и самоконтроля:
- Расскажите о цитоархитектонических полях коры полушарий больного мозга и расположении в коре ядер различных анализаторов.
- Назовите места расположения в коре полушарий большого мозга центров речи.
- Расскажите о строении и функциях лимбической системы и ретикулярной формации.
- Расскажите о классификации проводящих путей головного и спинного мозга.
- Укажите, в какие зоны коры полушарий большого мозга несут нервные импульсы проводящие пути болевой, температурной и других видов чувствительности.
- Расскажите, что вы знаете о строении и назначении пирамидных и экстрапирамидных проводящих путей.
Головной мозг. У новорожденного головной мозг относительно большой, масса его в среднем 390 г (340—430) у мальчиков и 355 г (330—370) у девочек, что составляет 12—13 % массы тела (у взрослого — примерно 2,5 %). К концу первого года жмзни масса головного мозга удваивается, а к 3—4 годам — утраивается. В дальнейшем (после 7 лет) масса головного мозга возрастает медленно и к
20—29 годам достигает максимального значения (1355 г — у мужчин и 1220 г — у женщин). В последующие возрастные периоды, вплоть до 60 лет у мужчин и 55 лет у женщин, масса мозга существенно не изменяется, а после 55—60 лет отмечается некоторое уменьшение ее.
У новорожденного лучше развиты филогенетически более старые отделы мозга. Масса ствола мозга равна 10,0—10,5 г, что составляет примерно 2,7 % массы тела (у взрослого — около 2%). К моменту рождения большинство ядер ствола мозга хорошо развито, отростки их нейронов миелинизированы.
Структуры среднего мозга к моменту рождения дифференцированы недостаточно. Такие ядра, как красное ядро, черное вещество, созревают в постнатальный период, формируя нисходящие проводящие пути экстрапирамидной системы.
Промежуточный мозг у новорожденного развит относительно хорошо. К моменту рождения дифференцированы специфические и неспецифические ядра таламуса, благодаря, чему сформированы все виды чувствительности. Становление ассоциативных ядер происходит после рождения и связано с развитием ассоциативных полей коры. Окончательное созревание таламических ядер заканчивается примерно к 13 годам.Структуры гипоталамуса у новорожденных недостаточно дифференцированы, в связи с чем у них несовершенны механизмы терморегуляции, регуляция обменных процессов. Дифференцировка ядер гипоталамуса происходит неравномерно. К 2—3-летнему возрасту большинство гипотало- мических ядер сформировано, но их окончательное функциональное созревание происходит к 15—16 годам.
Мозжечок. Масса мозжечка у новорожденного составляет 20 г (5,4 % массы мозга). К 5 мес жизни масса мозжечка увеличивается в 3 раза, к 9 мес — в 4 раза (ребенок умеет стоять, начинает ходить). У годовалого ребенка масса мозжечка составляет — 90 г. К 7 годам она достигает нижней границы массы мозжечка взрослого человека (130 г). Особенно интенсивное развитие структур мозжечка происходит в период полового созревания.
Полушария большого мозга. Новая кора в структурах полушарий начинает формироваться в конце второго месяца внутриутробного периода. На протяжении всей внутриутробной жизни в развитии неокортекса выделяют три периода: ранний миграционный; средний, или период предварительной дифференциров- ки на слои; поздний, или период заключительной дифференци- ровки. Ранний период охватывает промежуток со 2-го по 4-й лунный месяц. В это время наблюдается миграция нейробластов из глубоких (околожелудочковых) слоев конечного мозгового пузыря в корковую пластинку. В период с 7-й по 10-ю неделю
начинают формироваться нижние (глубокие) слои коры (V и VI). Несколько позже (на 13—15-й неделе) происходит диффе- ренцировка верхних слоев — I, II, III и IV. Начиная с 4-го месяца внутриутробной жизни происходит предварительная ци- тоархитектоническая дифференцировка коры на клеточные слои, образуются первичные борозды и извилины. На 5-м месяце внутриутробного периода появляются следующие первичные борозды: боковая, центральная, шпорная, борозда мозолистого тела. Вторичные борозды (лобные, височные и др.) начинают появляется с 6-го месяца внутриутробного периода. После
- го месяца внутриутробной жизни начинают формироваться третичные борозды. Появляются индивидуальные вариации рисунка борозд и извилин. Происходит значительное увеличение поверхности коры. К моменту рождения число нейронов достигает 14—16 млрд, как у взрослого человека. Нейроны в коре новорожденного имеют веретенообразную форму и слабое развитие дендритов. После рождения в течение первых трех лет происходит интенсивный рост отростков нейронов, их миели- низация, дифференцирование нейронов в слоях коры. В период от 3 до 10 лет увеличивается количество ассоциативных волокон. За счет интенсивного развития III слоя увеличивается толщина коры. В этот период в основном завершаются процессы развития корковых формаций. Однако тонкая дифференцировка в ассоциативных полях продолжается до 16—18 лет. Созревание различных корковых территорий идет асинхронно. Первыми дифференцируются поля соматосенсорной зоны коры, затем — двигательная кора, после чего формируются зрительная и слуховая проекционные корковые зоны. Ассоциативные поля коры вступают в процесс развития последними.
Базальные ядра в период внутриутробного развития созревают неравномерно. Бледный шар достаточно сформирован уже к моменту рождения. Хвостатое ядро и скорлупа чечевицеобразного ядра достаточно сформированными выглядят только в конце 1-го года после рождения. К семилетнему возрасту происходят окончательное созревание базальных ядер и формирование их связей с корой, что и обеспечивает выполнение более точных и координированных произвольных движений.
Вегетативная нервная система. У новорожденных симпатический и парасимпатический отделы вегетативной нервной системы сформированы недостаточно. Однако преобладает влияния симпатического отдела, которое сохраняется на протяжении 6—7 лет после рождения. По мере созревания структур мозга усиливается влияние вегетативной нервной системы на деятельность внутренних органов.
А так же в разделе «Проводящие пути головного и спинного мозга »
- Гипофиз, гормоны гипофиза
- Щитовидная железа
- Паращитовидные железы
- Надпочечники
- Эндокринная часть половых желез
- Эндокринная часть поджелудочной железы
- Шишковидное тело
- Одиночные гормонопродуцирующие клетки
- СЕРДЕЧНО-СОСУДИСТАЯ СИСТЕМА
- Работа сердца
- КРОВЕНОСНЫЕ СОСУДЫ ТЕЛА ЧЕЛОВЕКА
- Артерии большого круга кровообращения
- Вены большого круга кровообращения
- Возрастные особенности кровеносных сосудов
- Движение крови по сосудам
- Регуляция функций сердечно-сосудистой системы
- НЕРВНАЯ СИСТЕМА (ОБЩИЙ ПЛАН СТРОЕНИЯ И РАЗВИТИЯ НЕРВНОЙ СИСТЕМЫ)
- ФУНКЦИИ НЕРВНОЙ ТКАНИ
- ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА Спинной мозг
- Функции спинного мозга
- Головной мозг
- Мост
- Лимбическая система и ретикулярная формация
- Оболочки спинного и головного мозга
- Высшая нервная деятельность
- Качественные особенности высшей нервной деятельности человека
- Типы нервной деятельности
- Возрастные особенности высшей нервной деятельности человека