Пластическое звено метаболизма
Синтез мономеров и макромолекул, в том числе гормонов, ферментов, кофакторов является основным фактором жизнедеятельности клетки,
без которого невозможно предстваить нормальную жизнедеятельность организма.
Наиболее сложным и важным является процесс синтеза белка. От этого зависит приспособление к физиологическим потребностям при изменении внутренних и внешних условий. То есть синтез белка регулируется внешними и внутренними факторами и условиями, которые диктуют клетке какой набор белка и его количество необходимо синтезировать для выполнения физиологических функций.
Любая живая клетка способна синтезировать белки особенно в период роста и развития клеток (фаза клеточного цикла G1). В это время активно синтезируются белки для построения клеточных органоидов, мембран, синтезируются ферменты. Биосинтез белков идет интенсивно и в зрелых клетках, этим и определяется их функциональная активность: в клетках пищеварительных желез, синтезирующих белки-ферменты (пепсин, трипсин), в клетках желез внутренней секреции, синтезирующих белки- гормоны (инсулин, соматотропин), плазматические клетки синтезируют иммуноглобылины, Т-лимфоциты - цитокины и т.д.
Биосинтез белка - сложнейший многостадийный процесс синтеза полипептидной цепи в клетках живых организмов. Упрощенно биосинтез белка можно разделить на стадии транскрипции и трансляции.
Транскрипция - процесс считывания генетического кода с молекулы ДНК. В ДНК содержится и хранится информация о составе первичных структур разных белков. Отрезок ДНК, содержащий информацию о структуре одного белка, называют геном. Молекула ДНК представляет собрание множества генов. При этом на одной из цепочек ДНК синтезируется одноцепочечная молекула информационной или матричной РНК (мРНК). Каждой аминокислоте соответствует участок цепи ДНК из трех рядом состоящих нуклеотидов (кодон). Основную роль в транскрипции играет фермент РНК-полимераза.
Готовая мРНК, кодирующая аминокислотную последовательность будущей белковой цепи, образует затем сложный комплекс со специальной клеточной органеллой - рибосомой. На рибосомах идет второй этап биосинтеза белка - трансляция.
Трансляция заключается в синтезе полипептидной цепи в соответствии с информацией, закодированной в мРНК. Аминокислотная последовательность выстраивается при помощи транспортных РНК (тРНК). Каждой аминокислоте соответствует своя тРНК, имеющая соответствуюищий антикодон, «подходящий» к кодону мРНК. Во время трансляции рибосома движется вдоль мРНК, по мере этого наращивается полипептидная цепь. Энергией биосинотез белка обеспечивается за счет АТФ.
В дальнейщем при помощи вспомогательных белков шаперонов складывается биологически активная конформация пептидной цепи (свертывание). При посттрансляционном созревании у многих белков удаляют
ся части пептидной цепи или присоединяются дополнительные группы, например олигосахариды или липиды. Эти процессы происходят в эндо- плазматическом ретикулуме и в аппарате Гольджи. Затем готовая белковая молекула транспортируется в нужное место клетки.
Значительно проще синтезируются углеводы и липиды. Это ни что иное, как цикл простых биохимических реакций катализируемых ферментами. Исходными субстратами служат вещества, поступившие в клетку или полученные при метаболизме.
В условиях дефицита углеводов необходимая концентрация глюкозы в крови может поддерживаться за счет ее синтеза (глюконеогенез). Синтез глюкозы протекает, как и при гликолизе, но в обратном направлении. Исходными соединениями для глюконеогенеза являются некоторые аминокислоты, лактат, а также глицерин, то есть те вещества, которые способны превратиться в пируват или любой другой метаболит глюконеогенеза (ас- партат в оксалоацетат, глицерин в триозофосфат и т.д.). При различных физиологических состояниях для глюконеогенеза используются различные первичные вещества. В условиях голодания (недостаток углеводов получаемых с пищей) используется тканевой белок, который распадается до аминокислот. При интенсивной физической работе используется лактат, образующийся в эритроцитах и мышечной ткани при недостатке О2. При распаде жиров образуется глицерин. В организме человека за счет глюконео- генеза образуется несколько сотен граммов глюкозы в сутки.
Синтез высших жирных кислот может протекать в клетках различных органов и тканей, однако основная масса соединений этого класса синтезируется в печени и в жировой ткани, а важнейшим субстратом, продукты метаболизма которого используются для синтеза липидов, является глюкоза. С наибольшей интенсивностью этот синтез идет в период абсорбции глюкозы в желудочно-кишечном тракте, когда концентрация глюкозы в крови повышена.
Биосинтез липидов основан на синтезе жирных кислот из ацетил- КоА (образуется из глюкозы в результате окисления пирувата) с дальнейшим превращением их в жиры, воск, фосфолипиды и некоторые другие более специализированные биологически активные вещества. Для синтеза из ацетил-КоА в пальматиновую кислоту помимо ферментов (ацетил-СоА- карбоксилаза, пальмитилсинтетаза) требуется карнитин, осуществляющий перенос ацетил-КоА из митохондрий в цитоплазму, ацилпереносящий белок, на котором происходит сборка ацильных остатков и биотин кофер- мент ацетил-СоА-карбоксилазы. Под действием ферментов - элонгаз (удлинение цепи) и десатураз (введение двойных связей) протекает превращение пальматиновой кислоты в стеариновую и олеиновую.
Необходимо отметить, что ряд полиненасыщенных жирных кислот не синтезируется в организме, хотя они необходимы для нормального функционирования. Поэтому линолевая и линоленовая кислоты являются
незаменимыми (эссенциальными) и должны поступать в достаточном количестве с пищей. Арахидоновая кислота может синтезироваться в клетках животных из линоленовых кислот, однако в условиях недостаточного поступления линоленовой кислоты с пищей арахидоновая кислота также становится незаменимой жирной кислотой.
Ввообще все высшие жирные кислоты, всосавшиеся в клетки кишечника, используются в энтероцитах для ресинтеза различных липидов. При поступлении в энтероциты моноацилглицеринов, они через фосфатидную кислоту могут быть превращены в триацилглицерины. При поступлении в энтероциты лизофосфолипидов они превращаются в фосфолипиды.
Эндогенный синтез других липидов осуществляется в цитозоле клетки. Так для синтеза триглициридов и фосфолипидов необходим фосфоди- гидроксиацетон - промежуточный продукт расщепления глюкозы или высшие жирные кислоты и глицерин, поступающие в клетки из крови (рис. 50). Все необходимые организму глицерофосфолипиды могут синтезироваться в его клетках, причем в клетках могут функционировать несколько альтернативных метаболических путей биосинтеза глицерофосфолипидов.
Сфинголипиды, подобно глицерофосфолипидам, не являются незаменимыми компонентами пищи и могут синтезироваться из других соединений. Для их синтеза нужен в первую очередь сфингозин, активированные жирные кислоты в виде ацил-КоА-производных; активированный хо- лин или активированные мономеры углеводной природы в виде их УДФ- производных для синтеза цереброзидов или ганглиозидов.
Важное значение принадлежит и синтезу холестерина . Общее содержание холестерина в организме составляет около 140 г. Основная масса этого соединения включена в состав мембран всех клеток. Холестерин является предшественником в синтезе других стероидов: желчных кислот, стероидных гормонов, витамина D3. Существует два пути поступления холестерина (экзогенный и эндогенный). Суточная потребность человека в холестерине составляет около 1 г. Причем вся потребность в этом соединении может быть удовлетворена за счет его эндогенного синтеза. В то же время экзогенный, то есть пищевой холестерин также эффективно усваивается организмом.
Холестерин синтезируется в клетках из двух углеродных группировок ацетил-КоА (рис. 51). Процесс синтеза холестерина включает в себя порядка 35 последовательных реакций. Следует отметить, что некоторые промежуточные продукты этого метаболического пути используются для синтеза других соединений. Так, фарнезилпирофосфат используется в клетках для синтеза коэнзима Q, необходимого для работы главной дыхательной цепи митохондрий, или долихола, принимающего участие в синтезе гетероолигосахаридных компонентов гликопротеидов.
Все эти реакции требуют энергетических затрат. Энергия для синтеза доставляется реакцией расщепления АТФ. Поэтому каждое звено биосинтеза всегда сопряжено с распадом АТФ.
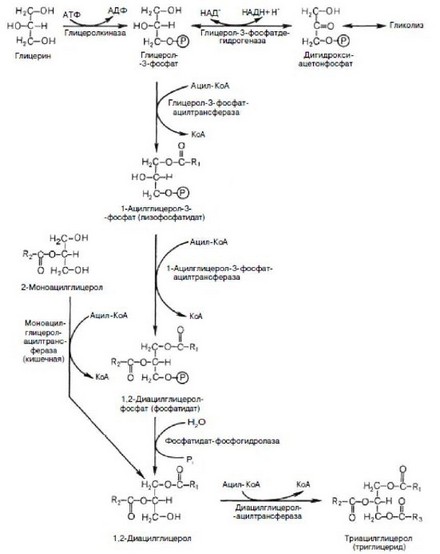
Рис. 50. Биосинтез триглицеридов (по Березову Т.Т., Коровкину Б.Ф.,
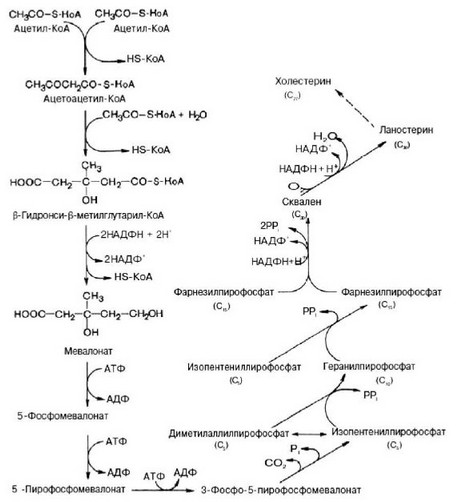
Рис. 51. Общая схема синтеза холестерина (по Березову Т.Т., Коровкину Б.Ф., 1998).
А так же в разделе «Пластическое звено метаболизма »
- Особенности метаболизма клетки в норме и при патологии
- Энергетическое звено метаболизма
- Утилизации продуктов метаболизма
- Метаболические механизмы реактивности клеток иммунной системыИзменение метаболизма в лимфоцитах в процессе их функционирования в норме и при патологии
- Метаболизм гранулоцитов и макрофагов в состоянии относительного покоя и при фагоцитозе
- Значение оксидоредуктаз в системе внутриклеточного метаболизма
- 2.4. Роль витаминов в системе клеточного метаболизма Межвитаминные взаимоотношения
- Глава 3 ВИТАМИНЫ И НАРУШЕНИЯ ФУНКЦИИ ИММУННОЙ СИСТЕМЫ
- Витаминная недостаточность и их клинические проявления
- Клиническая картина гиповитаминозов Гиповитаминоз A
- Гиповитаминоз В1
- Гиповитаминоз В2
- Гиповитаминоз В6
- Г иповитаминоз фолиевой кислоты (В9)
- Гиповитаминоз В12
- Гиповитаминоз C
- Гиповитаминоз D
- Гиповитаминоз E
- Недостаточность карнитина
- Лабораторные методы исследования витаминов Определение аскорбиновой кислоты в моче по Тильмансу
- Определение рибофлавина (витамина В2) в крови по Берчугу, Бессею и Лоури
- Определение рибофлавина (витамина В2) в моче по Е.М. Масленниковой и Л.Г. Г воздовой
- Определение 4-пиридоксиловой кислоты в моче по Хуффу и Перлцвейгу
- 3.4.8. Определение токоферола (витамина Е) в крови в модификации Фридемана