Активация В-лимфоцитов. Роль Т-клеток и цитокинов
С точки зрения роли в иммунологических процессах, В-лимфоциты являются клетками с двойственной функцией. С одной стороны, это анти- генраспознающие лимфоциты, способные дифференцироваться (обычно при участии Т-хелперов) в эффекторные клетки — антителопродуценты. С другой стороны, В-лимфоциты служат «профессиональными» АПК, способными активировать хелперые Т-лимфоциты. Схема их участия в гуморальном иммунном ответе, особенно при взаимодействии с Т-хелперами, очень сходна с аналогичной схемой участия макрофагов в клеточном ответе воспалительного типа.
Несмотря на то, что контакт В-лимфоцитов с патогенами и их продуктами возможен в очагах поступления патогенов в организм (в частности, в барьерных тканях), вовлечение этих клеток в первичный иммунный ответ возможно только во вторичных лимфоидных органах. Это обусловлено тем, что именно здесь создаются оптимальные условия для взаимодействия трех комопнен- тов пусковой реакции — антигена, наивной В-клетки и Т-хелпера типа Th2. Это взаимодействие происходит в межфолликулярном пространстве, или «короне» фолликула, где В- и Т-лимфоциты соседствуют друг с другом. Антиген доставляется в эти зоны (обычно с афферентной лимфой) не только в составе молекул MHC на поверхности дендритных клеток, но и в свободной форме.
Как было сказано выше, антигенсвязывающие рецепторы В-лимфоцитов — BCR — способны взаимодействовать как со свободным, так и мембраносвязанным антигеном в его нативной, нерасщепленной форме. Связывания антигена, самого по себе, недостаточно для активации В-клетки. Оно как бы обозначает те клоны В-лимфоцитов, которым предстоит участвовать в иммунном ответе. Для полноценной активации В-клетки требуется дополнительный сигнал, порождаемый контактным взаимодействием с Т-хелпе- ром (Th2).
При взаимодействии антигена с ВCR должен быть выполнен ряд условий, определяющих эффективность распознавания антигена. Прежде всего необходимо перекрестное сшивание иммуноглобулиновых рецепторов или иная форма их агрегации. Связывание антигеном двух и более рецепторов возможно при наличии в молекуле антигена повторяющихся эпитопов, что не характерно для белковых антигенов. Поэтому в действительности срабатывают другие, неспецифические механизмы агрегации мембранных рецепторов. Связывание антигена и агрегация BCR обусловливают кон- формационные изменения в мембранном иммуноглобулине. Эти изменения передаются на димер Iga/IgP (CD79a/b), а через него — на рецепторные киназы, которые при этом активируются. Важное усиливающее событие — взаимодействие антигена не только с мембранным IgM, но и с другими составляющими рецепторного комплекса. Доказан дополнительный вклад С3-компонента комплемента в активацию В-клеток антигеном (рис. 3.114). Образование комплекса антигена с мембранными рецепторами-антителами приводит к активации комплемента по классическому пути и расщеплению фактора С3 сначала до C3b, затем — до iC3b и C3d. Входящая в состав рецепторного комплекса В-клеток молекула CD21 является рецептором для комплемента (CR2), способным связывать фрагмент C3d. Возникающее при этом изменение конформации молекулы СD21 передается молекуле CD19 и вызывает активацию рецепторных тирозинкиназ, связанных с цитоплазматической частью CD19. Сочетанная активация тирозинкиназ, связанных с молекулами CD79 и CD19, формирует первый сигнал, ответственный за вовлечение в иммунный ответ клонов В-клеток, распознавших антиген.
За начальные этапы передачи сигнала о связывании антигена в В-клетках зответственны тирозинкиназы Lyn, Blk и Lck, фосфори- лирующие цитоплазматические части полипептидных цепей BCR и тирозинкиназу Syk. Этот фермент выполняет роль посредника между рецепторным аппаратом и внутриклеточными сигнальными факторами, подобно тирозинкиназе ZAP-70 Т-клеток, также относящейся к семейству Syk. Среди многочисленных молекул, активируемых Syk- киназой, важная роль принадлежит связанным с мембраной факторам, составляющим промежуточное звено передачи сигнала — адапторным и гуанозинсвязывающим (GEF) белкам, а также у-изоформе фосфолипазы С
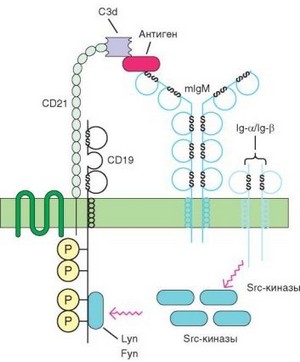
Рис. 3.114. Межмолекулярные взаимодействия в рецепторном комплексе В-клеток, необходимые для запуска активации и пролиферации В-клеток. Для активации В-клетки важны конформационные изменения мембранного иммуноглоублина в результате связывания антигена. Другое событие реализуется с участием системы комплемента. Образование комплекса антигена с мембранными антителами приводит к активации комплемента по классическому пути и расщеплению фактора С3 до C3d, распознаваемого молекулой CD21 (см. текст). За счет этих двух источников генерируется сигнал, ответственный за вовлечение в иммунный ответ конкретного клона В-клеток
(PLCy). Эти факторы обеспечивают дальнейшую передачу сигнала — уже по нескольким параллельным путям (рис. 3.115).
Однако для полноценной активации В-клеток, определяющей их дальнейшее развитие и дифференцировку в антителообразующие клетки, необходим дополнительный второй сигнал, возникающий в процессе контактного взаимодействия с Т-хелперами. Это взаимодействие, как уже отмечалось, происходит в морфологических структурах, где соседствуют Т- и В-лимфоциты — прежде всего в межфолликулярных пространствах. Как и в случае презентации антитена дендритными клетками Т-хелперам при контакте Т- и В-клеток, распознающих эпитопы одного и того же анти-
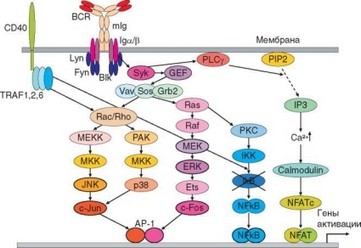
Рис. 3.115. Схема сигнальных путей при активации В-клеток. Схема в принципе близка таковой для активации Т-клеток (см. рис. 3.94). Основное отличие состоит в различной природе костимулирующего сигнала, поставляемого в В-клетку через молекулу CD40
гена, возникают сложности, обусловленные наличием малого числа клеток, специфичных к конкретному антигену. Это препятствие удается преодолеть лишь благодаря процессу непрерывной рециркуляции Т- и В-лимфоцитов.
Схематически процесс взаимодействия наивных В-клеток и ^2-лим- фоцитов представлен на рис. 3.116. После связывания антигена с BCR в В-клетке комплекс антиген—рецептор интернализуется, как это происходит в любой АПК. По ряду свойств, необходимых для презентации антигена (уровень эндоцитоза, эффективность процессинга поглощенных молекул антигенов и т.д.), В-лимфоциты значительно уступают дендритным клеткам. Однако они имеют существенное преимущество, которое состоит в избирательности поглощения антигенов при их распознавании BCR. Захваченный антиген процессируется и включается в состав молекул MHC-II так же, как это происходит в других АПК. Однако если в составе MHC на поверхности дендритной клетки на долю конкретного эпитопа может приходиться не более 0,1% комплексов пептид—MHC-II, то мембранные комплексы поверхности В-клетки существенно обогащены эпитопами тех антигенов, которые распознаются их рецепторами.
Принципиально важно, что в состав MHC-II избирательно включаются не те эпитопы, которые распознаются BCR (как известно, структурные характеристики В- и Т-эпитопов различны), а другие — Т-клеточные эпитопы. Выше уже рассматривалась природа В- и Т-эпитопов и их различия между собой (см. раздел 3.2). Практически каждая молекула имеет уникальный набор детерминант, распознаваемых рецепторами В- и Т-клеток. Если
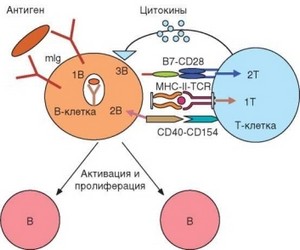
Рис. 3.116. Природа сигналов, генерируемых при взаимодействии В-клеток с Т-хелпе- рами и необходимых для активации и запуска пролиферации В-клеток. Связывание антигена с мембранным иммуноглобулином (mIg) В-клеточного рецептора служит источником 1-го активационного сигнала (1В), усиливающего экспрессию молекул MHC-II и костимулирующих молекул В7 (CD80 и CD86). Комплекс антиген-рецептор интернализуется, фрагменты антигена включаются в молекулы MHC-II, экспрессируемые на поверхности В-клетки. Т-клеточный рецептор ТИ2-клетки распознает комплекс MHC-II-пептид (индуцируется сигнал 1Т). Вместе с костимуляцией (сигнал 2Т) через CD28 это приводит к активации Т-клетки, экспрессии CD154 и секреции цитокинов. В результате взаимодействия CD40-CD154 возникает 2-й активационный сигнал для В-клеток (сигнал 2В), что приводит к экспрессии рецепторов для цитокинов. Цитокиновые сигналы, индуцируемые при связывании IL-2, IL-4, IL-5 с их рецепторами (3В), обеспечивают продвижение В-лимфоцитов по клеточному циклу и их пролиферацию
эти наборы и перекрываются, то только частично. В-лимфоцит распознает В-клеточные эпитопы своим ВCR и в то же время встраивает Т-клеточные эпитопы в состав молекул MHC-II, подготавливая лиганд для TCR. Эти лиганды распознают Т-клетки, принадлежащие к соответствующим клонам. Таким образом, в отношении В-эпитопов антигена В-клетка является распознающей, а в отношении Т-эпитопов — презентирующей.
На этом этапе иммунного процесса во взаимодействие с В-лимфоци- том вовлекаются не наивные Т-клетки (их могут активировать только
дендритные клетки), а ТИ2-лимфоциты, уже ранее получившие такой же антигенный сигнал от дендритных клеток в Т-зонах вторичных лимфоидных органов. Вовлечение во взаимодействие с В-лимфоцитами Th2-, а не TW-клеток, обусловлено направлением миграции этих типов Т-хелперов. Если TM-клетки из мест своего образования (Т-зоны) с эфферентным током лимфы попадают в рециркуляцию, то большинство ^2-клеток, экспрессирующих хемокиновый рецептор CXCR5, движется в сторону фолликулов и на этом пути встречается с В-лимфоцитами. В-лимфоцит и ^2-клетка взаимодействуют посредством иммунного синапса, принципиально аналогичного по своей структуре синапсу, формируемому между дендритной клеткой и Т-хелпером (см. раздел 3.4.1.3). Во время длительного существования синапса (часы) генерируются активационные сигналы, направленные в Т- и В-клетки (см. рис. 3.116). В ^2-клетки сигналы поступают через TCR и CD28, и это способствует экспрессии костимулирующей молекулы CD154 (лиганда молекулы CD40) и усилению секреции ^2-цитокинов. То и другое служит источником дополнительных сигналов для В-лимфоцита.
Принцип внутриклеточной передачи сигнала в В-лимфоцитах аналогичен таковому в Т-клетках. В активации В-клеток участвуют сходные сигнальные пути, приводящие к образованию тех же трех транскрипционных факторов — NF-AT, AP-1 NF-kB (см. раздел 3.5.2.1). Источником костимулирующих сигналов в случае В-клетки выступает молекула CD40, вовлекаемая в активацию в результате контактного взаимодействия с Т-хелпером. При связывании с ней CD40-лиганда (CD154) генерируется костимулирующий сигнал, в проведении которого основную роль играют молекулы TRAF1, TRAF2, и в меньшей степени TRAF6. Этот сигнал играет важную роль в формировании Rac-зависимых ветвей MAP-каскада, приводящих к активации киназ JNK и р38. Эти киназы нужны для экспрессии белка c-Jun, участвующего в формировании транскрипционного фактора АР-1. CD40/TRAF-зависимый путь необходим также для активации фосфатазы IKK, обеспечивающей дефосфорилирование ингибиторной субъединицы IkB, что приводит к ее расщеплению и активации транскрипционного фактора NF-kB. Подобно процессу активации Т-клеток, наименее зависим от костимуляции при активации В-клеток Са2+-зависимый путь, приводящий к формированию транскрипционного фактора NF-AT (см. рис. 3.115).
Последствия активации В-клеток аналогичны таковым при активации Т-лимфоцитов (см раздел 3.5.2.2). Они состоят в экспресси комплекса генов, необходимых для реализации первого этапа реакции клонов, вовлекаемых в иммунный ответ, — их пролиферативной экспансии. Для В-клеток это означает прежде всего появление на их поверхности рецепторов для цитокинов, которые обеспечивают сначала их пролиферацию, а затем дифференцировку: IL-4, IL-5, IL-6, IL-10, IL-1, IL-2. В отличие от Т-хелперов, В-лимфоциты не обеспечивают себя этими цитокинами, а зависят в этом отношении от других клеток. Ситуация облегчается тем, что почти все эти цитокины секретируются ^2-лимфоцитами, с которыми контактируют В-клетки. IL-2, не вырабатываемый ^2-клетками, присутствует в зоне активации в достаточном количестве. Его секретируют CD4+ Т-клетки (в том числе предшественники Th2-лимфоцитов), активируемые дендритными клетками. Источником IL-1 служат главным образом макрофаги.
Активацию и последующее развитие В-лимфоцитов контролирует целый комплекс цитокинов. Рецепторы для них появляются на поверхности В-клеток во время фазы G1 клеточного цикла. Среди факторов, индуцирующих пролиферацию активированных В-клеток, раньше других (в фазу G1a) свою активность проявляют IL-4 (основной ростовой фактор) и IL-1 (кофактор). Уже после преодоления точки рестрикции (в фазе G1b) основными факторами, обусловливающими продвижение клетки по циклу, становятся IL-2 и IL-5, при участии которых В-клетка вступает в митоз. Как правило, на активированную В-клетку одновременно действует не один, а несколько цитокинов, среди которых основной — IL-4. Из сказанного следует, что цитокины, участвующие в индукции пролиферации В-клеток, относятся к разряду заменимых и мутации генов каждого из них не сказываются на пролиферативной экспансии клонов В-лимфоцитов.
После завершения митоза клетки не возвращаются в фазу покоя (G0); для продолжения пролиферации им нужны сигналы от цитокинов, но не контакт с Т-лимфоцитами. Как и Т-клетки, стимулированные В-лимфоциты совершают 6—8 делений за 5—7 сут (продолжительность периода пролиферации определяется длительностью экспрессии рецепторов на поверхности В-клеток). Одновременно с делением происходит реализация дифференцировочной программы, запускаемой при активации В-клеток. Завершение этой программы в значительной степени зависит от микроокружения В-клеток.
Источник: Ярилин.А.А , «Иммунология » 2010
А так же в разделе «Активация В-лимфоцитов. Роль Т-клеток и цитокинов »
- Презентация антигена
- Миграция клеток, участвующих в презентации антигена
- Иммунный синапс
- Костимуляция
- Активация Т-лимфоцитов
- Пролиферативная экспансия клонов Т-хелперов
- Дифференцировка Т-хелперов
- Th17 и другие адаптивные субпопуляции Т-клеток
- ИММУННЫЙ ОТВЕТ
- Клеточный иммунный ответ
- Воспалительный Т-клеточный иммунный ответ
- Гуморальный иммунный ответ
- Дифференцировка и селекция В-клеток в зародышевых центрах
- Гибридомы и моноклональные антитела. Генно-инженерные антитела
- Иммунологическая память и вторичный иммунный ответ
- В-клетки памяти
- Т-клетки памяти
- Вторичный иммунный ответ
- Неклассические проявления иммунных реакций
- 3.6.4.3. Проявления активности у5Т- и CD8aa+ Т-клеток
- Иммунологические функции NKT-клеток
- Иммунные процессы в слизистых оболочках (мукозальный иммунный ответ)
- Афферентное и центральное звенья мукозального иммунного ответа
- Роль миграции клеток в мукозальном иммунитете
- Эффекторные механизмы мукозального иммунитета
- Контроль и регуляция иммунного ответа
- Генетический контроль иммунного ответа
- Эндокринный и нервный контроль иммунного ответа
- Регуляция иммунного ответа
- Регуляторные Т-клетки
- РЕЗЮМЕ