Эффекторные механизмы мукозального иммунитета
Поступившие из кровотока CD4+ Т-клетки задерживаются в подслизистом слое и мигрируют из него в lamina propria и (в небольшом количестве) в эпителиальный слой. CD8+ Т-клетки, напротив, мигрируют преимущественно в эпителиальный слой слизистых оболочек и пополняют пул внутриэпителиальных лимфоцитов.
Свойства клеток, мигрирующих в лимфоидные ткани, ассоциированные со слизистыми оболочками, могут изменяться под влиянием микроокружения последних. Это происходит уже в процессе трансмиграции через эндотелий сосудов. Дополнительный эффект оказывает взаимодействие с местными АПК. Наконец, на мигрирующие клетки сильно влияют гуморальные факторы микроокружения, прежде всего цитокины. Так, при миграции в слизистые оболочки (особенно в респираторном тракте) Т-хелперы предпочтительно дифференцируются в хелперы Th2-rarn. Даже уже сформировавшиеся Thl-хелперы могут перестраивать свою дифференцировочную программу и превращаться в ^2-клетки. Этому способствует прежде всего наличие в микроокружении IL-4 — основного фактора, определяющего дифференцировку ^2-клеток, секретируемого тучными клетками laminae propriae. Имеет значение также высокая экспрессия на АПК слизистых оболочек дыхательных путей костимулирующей молекулы ICOS, запускающей в Т-клетках сигнальный путь, который поддерживает дифференцировку в ^2-лимфоциты.
Главный продукт гуморального иммунного ответа на начальных этапах его развития (4-7-е сутки) — IgM-антитела, поступающие в системный кровоток и не играющие основной роли в защите слизистых оболочек. Однако уже в ходе первичного иммунного ответа происходит переключение изотипов иммуноглобулинов. В микроокружении слизистых оболочек мигрирующие сюда и образовавшиеся местно плазматические клетки переключают изотип секретируемых антител на IgA. Пик IgA-ответа в слизистых оболочках дыхательных путей приходится на 7—10-е сутки иммунного ответа. Среди секретируемых IgA-антител преобладают молекулы изотипа IgA1 (рис. 3.131). Избирательное переключение изотипа антител на IgA характерно для участков слизистых оболочек, заселенных микроорганизмами (так, в миндалинах и в кишечнике человека на долю IgA-образующих клеток приходится до 90% антителопродуцентов). В свободных от микрофлоры слизистых оболочках (например, нижних дыхательных путей) преобладают IgG-продуценты.
Важнейший эффекторный фактор мукозальных лимфоидных тканей — секреторные IgA, формируемые из обычных димерных молекул IgA при
Местная дифференцировка в мукозальной ткани
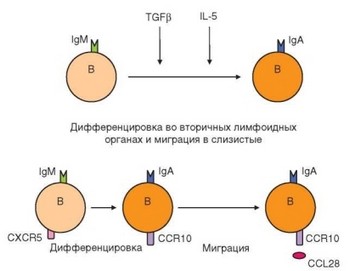
Рис. 3.131. Продуценты IgA-антител в слизистых оболочках. IgA образуется в мукозальном отделе иммунной системы как местно (под влиянием локальных факторов), так и во вторичных лимфоидных органах, откуда они мигрируют в слизистые оболочки
транспорте через эпителиальный слой слизистых оболочек (рис. 3.132). На базальной поверхности эпителиальных клеток слизистых оболочек присутствуют так называемые полимерные иммуноглобулиновые рецепторы ^IgR), способные взаимодействовать с димерными молекулами IgA и в меньшей степени — с пентамерами IgM. После связывания образовавшийся комплекс интернализуется эпителиальной клеткой и транспортируется в составе везикулы от базального к апикальному концу клетки. В процессе транспортировки происходит отщепление большей части рецептора, которая в виде секреторного компонента (SC, молекулярная масса 60 кДа) становится составной частью молекулы секреторного IgA (sIgA). При достижении апикальной поверхности клетки содержимое везикулы, включая sIgA, выбрасывается в просвет органа. Присутствие SC-цепи в составе sIgA придает молекуле устойчивость к действию протеаз, присутствующих в среде (особенно в пищеварительном тракте). IgA не являются ни опсонизирующим фактором, ни активатором комплемента. Их защитная функция проявляется иначе: связываясь с микроорганизмами-мишенями, IgA-антитела ослабляет их подвижность, предотвращают адгезию на эпителиальных клетках и, как следствие, — проникновение в обход М-клеток. С другой стороны, образование комплекса sIgA с антигенами возбудителей облегчает поглощение последних М-клетками.
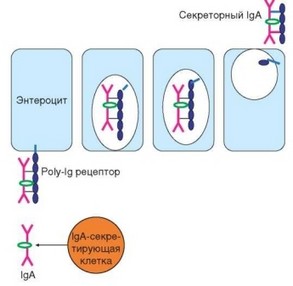
Рис. 3.132. Механизм формирования секреторного IgA. IgA, секретируемый мукозальными плазматическими клетками, взаимодействует с поли-^-рецептором; комплекс интернализуется и транспортируется в апикальную часть клетки. После протеолиза значительная часть поли-^-рецептора отделяется от мембраны и в форме секреторного компонента входит в состав секреторного IgA, который поступает в просвет кишечника (или других полых органов) по механизму экзоцитоза
Клеточные механизмы адаптивного иммунитета вносят в иммунную защиту слизистых оболочек существенный вклад. В первую очередь это относится к цитотоксическим Т-лимфоцитам, играющим основную роль в противовирусной защите слизистых оболочек, особенно в воздухоносных путях. CD8+ Т-клетки эпителия и собственной пластины убивают инфицированные вирусом клетки, обеспечивая тем самым защиту от вируса гриппа и других респираторных вирусов. TM-клетки усиливают воспалительный ответ макрофагов: они стимулируют фагоцитарную и бактерицидную активность преимущественно через выработку IFNy, что обеспечивает защиту от патогенов, локализующихся во внутриклеточных гранулах.
Как и при осуществлении защиты первой линии, при реализации иммунного ответа в слизистых оболочках большую роль играют регуляторные клетки, которые сдерживают иммунные процессы, предотвращая их деструктивные проявления и развитие аутоагрессии. Пул этих клеток пополняется индуцибельными (адаптивными) регуляторными Т-клетками, к которым относят регуляторные T-клетки 1-го типа (Tr1), продуцирующие IL-10, и Th3, секретирующие TGFp. Таким образом, в защите барьерных тканей от «иммунного повреждения» участвуют регуляторные механизмы как врожденного, так и адаптивного иммунитета.
- Развитие мукозального иммунного ответа при повторном контакте с патогеном
При развитии мукозального иммунного ответа формируются клетки памяти, избирательно мигрирующие в барьерные ткани (особенно в те, в которых они образовались). CD8+ Т-клетки памяти локализуются в эпителиальном слое; CD4+ Т- и В-клетки памяти — преимущественно в подслизистом слое, а также в структурированных лимфоидных образованиях. В большинстве отделов лимфоидных тканей слизистых оболочек преобладают В-клетки памяти, экспрессирующие мембранный рецептор IgA-изотипа, но в бронхолегочном отделе — IgG-изотипа. При иммунном ответе Т-клетки памяти могут попадать в орган, например, при инфицировании респираторными вирусами.
При повторном поступлении в организм патогена через слизистые оболочки он взаимодействует с иммунной системой, обогащенной клетками памяти. Как известно, презентация антигена Т-клеткам памяти значительно облегчена и не требует вовлечения всех механизмов, необходимых для его презентации наивным Т-клеткам. Именно поэтому антиген Т-клеткам памяти могут презентировать не только дендритные клетки, но и макрофаги, В-лимфоциты и другие клетки, возможно, даже эпителиальные. Таким образом, региональный лимфатический узел — не единственное и даже не основное место, где может происходить запуск вторичного иммунного ответа. Особое значение при этом приобретают события, происходящие в «кармане» М-клеток, где соседствуют Т- и В-клетки памяти. В-лимфоциты памяти выступают в этой ситуации в качестве АПК. За этим следует активация Т-клеток памяти и их дифференцировка в эффекторные лимфоциты. При вторичном, как и при первичном иммунном ответе, дифференцировка СD4+ Т-клеток осуществляется главным образом в направлении ^2-лим- фоцитов, что способствует преимущественному развитию гуморального иммунного ответа. Активированные in situ В-клетки памяти также могут получать помощь от ^2-клеток непосредственно в структурах мукозальных лимфоидных тканей с преобладающим образованием IgA-продуцентов. Таким образом, вторичный иммунный ответ в слизистых оболочках может реализоваться «местными» клетками без значительной их миграции извне и его интенсивность существенно выше, чем первичного.
Источник: Ярилин.А.А , «Иммунология » 2010
А так же в разделе «Эффекторные механизмы мукозального иммунитета »
- Презентация антигена
- Миграция клеток, участвующих в презентации антигена
- Иммунный синапс
- Костимуляция
- Активация Т-лимфоцитов
- Пролиферативная экспансия клонов Т-хелперов
- Дифференцировка Т-хелперов
- Th17 и другие адаптивные субпопуляции Т-клеток
- ИММУННЫЙ ОТВЕТ
- Клеточный иммунный ответ
- Воспалительный Т-клеточный иммунный ответ
- Гуморальный иммунный ответ
- Активация В-лимфоцитов. Роль Т-клеток и цитокинов
- Дифференцировка и селекция В-клеток в зародышевых центрах
- Гибридомы и моноклональные антитела. Генно-инженерные антитела
- Иммунологическая память и вторичный иммунный ответ
- В-клетки памяти
- Т-клетки памяти
- Вторичный иммунный ответ
- Неклассические проявления иммунных реакций
- 3.6.4.3. Проявления активности у5Т- и CD8aa+ Т-клеток
- Иммунологические функции NKT-клеток
- Иммунные процессы в слизистых оболочках (мукозальный иммунный ответ)
- Афферентное и центральное звенья мукозального иммунного ответа
- Роль миграции клеток в мукозальном иммунитете
- Контроль и регуляция иммунного ответа
- Генетический контроль иммунного ответа
- Эндокринный и нервный контроль иммунного ответа
- Регуляция иммунного ответа
- Регуляторные Т-клетки
- РЕЗЮМЕ