Миграция клеток, участвующих в презентации антигена
Взаимодействуя в барьерных тканях с патогенами — носителями антигенов, дендритные клетки поглощают их с помощью различных форм эндоцитоза и под влиянием провоспалительных цитокинов мигрируют в тканевую жидкость, а затем — в лимфу, где они приобретают характерную форму вуалевых клеток (рис. 3.83). Одновременно изменяется мембранный фенотип этих клеток: усиливается экспрессия молекул MHC-II, костимулирующих молекул, появляются ргинтегрины и хемокиновый рецептор CCR7. В это же время осуществляется процессинг антигена и экспрессия его пептидов на поверхности клетки в составе молекул MHC. С током афферентной лимфы дендритные клетки проникают в региональные лимфатические узлы через их выпуклую поверхность, противоположную воротам. Попав в ткань лимфатического узла, дендритные клетки мигрируют в Т-зоны, куда их привлекают хемокины CCL19 (ELC) и CCR21 (SLC), распознаваемые рецептором CCR7 (рис. 3.84). Эти хемокины секретируются стромальными клетками Т-зон лимфатического узла. При вовлечении регионального лимфатического узла в воспалительный процесс (что обычно происходит при локальном инфицировании) проникновению дендритных клеток в лимфатический узел способствуют также вырабатываемые в нем провоспалительные хемокины CCL2, CCL7, CCL8 (МСР1, МСР2 и МСР3). Оказавшись в Т-зоне лимфоидных органов, дендритные клетки созревают (признак созревания — экспрессия молекул CD83) и превращаются в интердигитальные клетки. Эти клетки не экспрессируют рецептор CCR7, но сами вырабатывают хемокины CCL21 и CCL19. Таким образом, они сами начинают привлекать как незрелые дендритные клетки, пополняющие
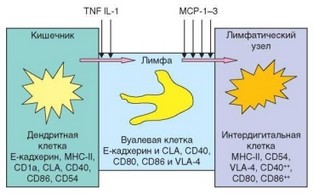
Рис. 3.83. Созревание дендритных клеток и их миграция из барьерных тканей в лимфатические узлы. Показано не только перемещение дендритных клеток, но и их созревание с приобретением способности презентировать антигенный пептид Т-лимфоцитам
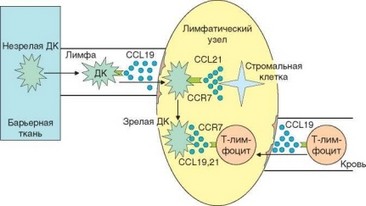
Рис. 3.84. Миграция дендритных клеток и Т-лимфоцитов в Т-зоны лимфатического узла определяет возможность контакта этих клеток. Для эффективной презентации антигена дендритные клетки и рециркулирующие Т-лимфоциты, поступающие в региональный лимфоузел разными путями, должны оказаться в одной его морфологической зоне (Т-зоне). Это достигается благодаря экспрессии клетками обоих типов рецептора CCR7, который распознает хемокины CCL19 и CCL21, секретируемые клетками высокого эндотелия, а также стромальными клетками (в том числе дендритны- ми/интердигитальными) Т-зон
популяцию интердигитальных клеток лимфоузлов, так и Т-лимфоциты, поступающие в узел в процессе рециркуляции. Это способствует сближению дендритных клеток с Т-лимфоцитами, необходимому для формирования иммунного синапса.
Накопление Т-клеток необходимой специфичности в региональном лимфатическом узле происходит с участием специального механизма. Как было детально описано выше (см. раздел 3.4.2.5), Т-лимфоциты непрерывно рециркулируют, при этом они периодически поступают в лимфоидные органы, прежде всего в лимфатические узлы. Т-клетки проникают в лимфоузлы с током крови (т.е. через ворота органа — иным путем, чем дендритные клетки) и мигрируют в ткань узла через высокий эндотелий посткапиллярных венул. Экстравазация происходит с участием экспрессированного на Т-лимфоцитах L-селектина CD62L (рецептора хоминга) и хемокинового рецептора лимфоцитов CСR7 (распознает хемокин CCL21, секретируемый эндотелиальными клетками). Затем Т-клетки мигрируют в Т-зоны по градиенту хемокинов CCL19 и CCL21, вырабатываемых, как указано выше, стромальными клетками Т-зон и локализованными здесь интердигитальными клетками. Обычно Т-лимфоциты не задерживаются в лимфатическом узле долго и, покидая его с лимфой, вступают в очередной цикл рециркуляции.
Ситуация складывается по-иному в отношении тех Т-клеток, чьи анти- генраспознающие рецепторы специфичны для эпитопов, представлен-
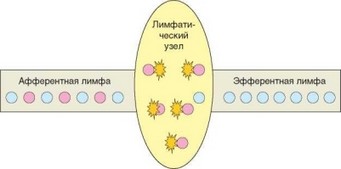
Рис. 3.85. Улавливание специфических клонов Т-лимфоцитов в региональном лимфатическом узле. Проблема недостатка Т-клеток любого конкретного клона в лимфатическом узле решается путем улавливания этих клеток в процессе их рециркуляции через лимфоузел
ных на поверхности интердигитальных клеток в комплексе с молекулами МНС. В Т-зонах лимфоидных органов, в частности лимфатических узлов, Т-лимфоциты и интердигитальные клетки находятся в достаточно тесном контакте и непрерывно взаимодействуют друг с другом с помощью молекул адгезии и различных мембранных рецепторов. При наличии сродства между TCR Т-клетки и комплексом MHC—пептид интердигитальной клетки формируется зона устойчивого межклеточного контакта — иммунный синапс. Процесс рециркуляции непрерывен, и через каждый узел может пройти любая Т-клетка, в том числе Т-клетки клона, специфичного к антигену, поступившему в организм. Поэтому практически все Т-лимфо- циты, принадлежащие тому клону, который специфичен к комплексам пептид-МНС, присутствующим на поверхности интердигитальных клеток, взаимодейсвуют с дендритными клетками регионального лимфатического узла и задерживаются в нем. При этом эфферентная лимфа оказывается обедненной Т-клетками клонов, вовлекаемых в иммунный ответ (рис. 3.85). Описанный процесс называется улавливанием (рекрутированием) клонов Т-лимфоцитов. С его помощью решается первая из двух упомянутых выше проблем — проблема дефицита антигенспецифических Т-клеток. Значимость этого процесса можно проиллюстрировать простым опытом: удаление регионального лимфатического узла через несколько часов после иммунизации мышей антигеном с стимулятором-адъювантом приводит к утрате способности животных к иммунному ответу на данный антиген, т.к. вместе с лимфатическим узлом из организма удаляются задержанные в нем Т-клетки специфического клона.
Источник: Ярилин.А.А , «Иммунология » 2010
А так же в разделе «Миграция клеток, участвующих в презентации антигена »
- Презентация антигена
- Иммунный синапс
- Костимуляция
- Активация Т-лимфоцитов
- Пролиферативная экспансия клонов Т-хелперов
- Дифференцировка Т-хелперов
- Th17 и другие адаптивные субпопуляции Т-клеток
- ИММУННЫЙ ОТВЕТ
- Клеточный иммунный ответ
- Воспалительный Т-клеточный иммунный ответ
- Гуморальный иммунный ответ
- Активация В-лимфоцитов. Роль Т-клеток и цитокинов
- Дифференцировка и селекция В-клеток в зародышевых центрах
- Гибридомы и моноклональные антитела. Генно-инженерные антитела
- Иммунологическая память и вторичный иммунный ответ
- В-клетки памяти
- Т-клетки памяти
- Вторичный иммунный ответ
- Неклассические проявления иммунных реакций
- 3.6.4.3. Проявления активности у5Т- и CD8aa+ Т-клеток
- Иммунологические функции NKT-клеток
- Иммунные процессы в слизистых оболочках (мукозальный иммунный ответ)
- Афферентное и центральное звенья мукозального иммунного ответа
- Роль миграции клеток в мукозальном иммунитете
- Эффекторные механизмы мукозального иммунитета
- Контроль и регуляция иммунного ответа
- Генетический контроль иммунного ответа
- Эндокринный и нервный контроль иммунного ответа
- Регуляция иммунного ответа
- Регуляторные Т-клетки
- РЕЗЮМЕ