Костимуляция
Помимо антигенраспознающего рецептора и корецепторов на поверхности Т-лимфоцита присутствуют костимулирующие молекулы. Если коре- цептор служит как бы дополнением рецептора и действует с ним как единой целое, запуская определенные сигнальные пути, то костимулиру- ющие молекулы действуют независимо, хотя их действие в конечном счете направлено на усиление сигнала, поставляемого рецептором/корецептором. На поверхности АПК также представлены костимулирующие молекулы, которые взаимодействуют с костимулирующими молекулами Т-клеток. Костимуляция Т-клеток — обязательный компонент презентации антигена и условие их эффективной активации (рис. 3.88). Презентация антигена без костимуляции приводит к развитию анергии Т-клеток.
Известно несколько пар костимулирующих молекул, участвующих в презентации антигена (табл. 3.24, рис. 3.89). Одна из молекул каждой пары представлена на поверхности Т-лимфоцита, а другая экспрессируется АПК. Взаимное распознавание этих молекул служит источником вспомогательных сигналов, имеющих определенную направленность — чаще от дендритной клетки к Т-хелперу (для усиления сигнала, поступающего от TCR), но иногда — от Т-лимфоцита к АПК. Система костимулирующих молекул строго организована и имеет ряд характерных особенностей. Так, в каждой паре взаимодействующих молекул одна является конститутивной, т.е. спонтанно экспрессируется на покоящихся клетках, а вторая индуцируется при
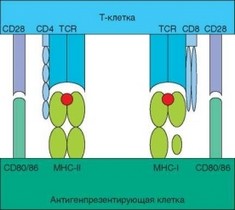
Рис. 3.88. Схема презентации антигена. Представлены все основные пары молекул, участвующие в презентации — молекулы MHC, несущие антигенный пептид, и TCR, распознающий их с участием корецепторов (CD4, CD8), а также костимулирующие молекулы В7 (CD80/CD86) и CD28
активации клетки (иногда это экспрессия de novo, иногда усиление исходно слабой экспрессии). Сигналом для экспрессии зачастую служит взаимодействие другой пары костимулирующих молекул. Для некоторых пар костимулирующих молекул характерна избыточность: костимулирующую функцию выполняет не одна, а две или более молекул со сходными, хотя и не полностью идентичными функциями. Наконец, на одной клетке могут экспрессироваться лиганды одних и тех же костимулирующих молекул, включающие противоположные по эффекту (усиливающий и супрессорный) сигналы.
Таблица 3.24. Костимулирующие молекулы, участвующие в презентации антигена
|
Название |
Семейство, молекулярная масса |
Локализация на клетках |
Лиганд (на АПК) |
Сигнальный мотив / связывающий мотив лиганда |
Эффект от передачи сигнала в клетку |
|
CD28 |
Суперсемейство иммуноглобулинов, 44 кДа |
Т-клетки |
B7-1 (CD80), B7-2 (CD86) |
YXXM/ MYPPPY |
Костимуляция, секреция IL-2 |
|
CTLA-4 |
Суперсемейство иммуноглобулинов, 33—37 кДа |
Активиро ванные Т-клетки |
B7-1 (CD80), B7-2 (CD86) |
YXXM/ MYPPPY |
Подавление активности Т-клеток |
|
ICOS |
Суперсе мейство иммуног лобулинов |
Активиро ванные Т-клетки (Th2gt;Th1), NK-клетки |
ICOSL |
YXXM/ FDPPPF |
Костимуляция, секреция цитокинов (Th2gt;Th1) |
|
PD-1 |
Суперсе мейство иммуног лобулинов |
Активиро ванные Т-клетки, активированные B-клетки, макрофаги |
PD-L1 |
ITIM |
Подавление развития и активности Т-клеток |
|
BTLA |
Суперсе мейство иммуног лобулинов |
Активиро ванные Т-клетки (Th1gt;Th2), В-клетки |
B7-H4 |
2 ITIM |
Подавление активации Т-клеток (Th1gt;Th2) |
|
CD154 (CD40L) |
TNF, 34-39 кДа |
Активированные Т-клетки, NK-клетки, B-клетки, тучные клетки, базофилы и эозинофилы |
CD40 |
Нет данных |
Костимуляция |
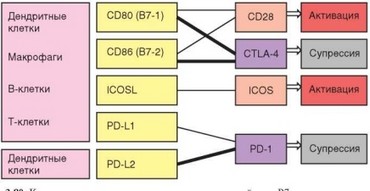
Наиболее известная и важная система костимуляции — передача в Т-хелпер сигнала через молекулу CD28, в результате ее взаимодействия с молекулами CD80 и/или CD86, расположенными на поверхности АПК. CD28 — гомодимерная трансмембранная молекула. Внеклеточная часть каждой цепи содержит домен суперсемейства иммуноглобулинов (V-домен), а также спейсерный (соединительный) участок. В ее внутриклеточной части есть сайт взаимодействия с липидной киназой PI3K. CD28 спонтанно экспрессируется на большинстве (около 80%) Т-клеток. Лиганды молекулы CD28 — две сходные по структуре молекулы — CD80 (В7-1) и CD86 (В7-2). Эти молекулы имеют между собой высокую гомологию и образованы двумя доменами суперсемейства иммуноглобулинов (один — V-, другой — С-типа). Они не экспрессируются (CD80) или слабо экспрессируются (CD86) на поверхности покоящихся дендритных и других АПК. Сигналом к индукции или усилению их экспрессии служит взаимодействие другой пары костимулиру- ющим молекул — CD40—CD154, которая будет описана ниже. Максимальную экспрессию молекулы CD86 наблюдают через 2 сут, а CD80 — через 4—5 сут после иммунизации, из чего следует, что молекула CD86 в большей степени, чем CD80, отвечает за костимуляцию на ранних этапах презентации антигена. Взаимодействие CD28 и CD80/86 происходит за счет взаимного связывания N-концевых частей их V-подобных доменов с участием мотива MYPPPY молекулы CD28. В результате взаимодействия происходит усиление активирующего сигнала, исходящего от рецепторного комплекса TCR—CD3, что и обозначают термином «костимуляция». Через CD28 в Т-клетки поступают сигналы, необходимые для поддержания их жизнеспособности, усиления адгезии и выработки цитокинов, особенно IL-2. Участие молекулы CD28 в презентации антигена сокращает число взаимодействий TCR—(MHC—пептид), необходимых для формирования активационного сигнала. При этом сокращается длительность межклеточного контакта. Как уже упоминалось, в отсутствие костимуляции передача сигнала через рецептор не только не
приводит к активации Т-клеток, но и вызывает противоположный эффект — клеточную анергию, т.е. неспособность Т-клеток и в последующем отвечать на сигналы, поставляемые через TCR. Молекулярные механизмы костиму- ляции будут рассмотрены ниже (см. раздел 3.4.2.1).
Взаимодействие CD28 с CD80/86 служит сигналом к экспрессии другого лиганда CD80/86 — молекулы CTLA-4 (CD152) (см. рис. 3.89), название который означает: «молекула 1 активации цитотоксических лимфоцитов» (Cytotoxic T lymphocyte activation molecule 1), что связано с первоначальным обнаружением ее на активированных цитотоксических Т-клетках. Т-хелпе- ры экспрессируют CTLA-4 вскоре после начала костимуляции, но вначале эта молекула находится внутри клетки и только через 48—72 ч появляется на ее поверхности в зоне иммунного синапса. Молекулы CTLA-4 и CD28 гомологичны (30% гомологии), но CTLA-4 обладает в 1000—2500 раз более высоким сродством к CD80/86, причем его специфичность также обусловлена мотивом MYPPPY. Главная особенность связывания CTLA-4 состоит в том, что он поставляет не костимулирующий, а ингибирующий сигнал. Учитывая более высокое сродство CTLA-4, чем CD28 к CD80/86, становится очевидно, что функция CTLA-4 состоит в завершении цепи активационных событий. Экспрессируемая при активации Т-клетки молекула CTLA-4 вовлекается в иммунный синапс, и осуществляемая через нее передача сигнала служит последним событием, реализуемым в иммунном синапсе, после чего он прекращает свое существование.
Описаны дополнительные пути костимуляции, обусловленные взаимодействием пар молекул ICOS (Inducible costimulator — индуцибельный костимулятор) и ICOS-L (L — лиганд), а также OX40 и OX40L. В обеих парах на первом месте указана молекула-рецептор, экспрессируемая Т-хелпером, а на втором — ее лиганд, экспрессируемый на АПК. Молекулы ICOS и OX40 передают в Т-клетку костимулирующие сигналы. Супрессорные аналоги костимулирующих молекул участвуют в регуляции активности Т-клеток и индукции толерантности.
Другую группу костимулирующих молекул образуют мембранные молекулы CD40 и CD40L (CD154). CD40 конститутивно экспрессируется на АПК. Эту молекулу относят к семейству рецепторов фактора некроза опухоли (TNF) (см. табл. 2.30). Она имеет 4 внеклеточных домена с 6 остатками цистеина, но не содержит домена смерти, характерного для многих представителей этого семейства. CD154 — трансмембранная молекула типа II (ее N-конец направлен внутрь клетки). Внеклеточный домен этой молекулы принадлежит к семейству TNF: он образован 8 антипараллельными р-сло- ями («гелевый рулет»). CD154 экспрессируют активированные Т-клетки, как правило, на 3-4-е сутки иммунного ответа. При взаимодействии CD40 с CD154 происходит тримеризация CD40, что необходимо для передачи сигнала внутрь клетки. Особенность этой пары костимулирующих молекул состоит в том, что сигнал от их взаимодействия направлен преимущественно или исключительно в сторону АПК, а не Т-лимфоцита.
Через взаимодействие CD40 с CD40L происходит активация дендритных клеток, макрофагов и В-лимфоцитов. Активация через CD40 усиливает экспрессию молекул, участвующих в презентации антигена, и стимулирует выработку цитокинов, необходимых для активации Т-клеток. Собственно хелперная функция Т-клеток в отношении В-лимфоцитов, необходимая для осуществления гуморального иммунного ответа, реализуется именно через этот путь костимуляции.
Помимо передачи сигналов через антигенраспознающий рецептор, корецеп- торы и костимулирующие молекулы, для активации Т-клеток важны сигналы, поступающие через молекулы интегринов (LFA-1, VLA-4) и другие молекулы адгезии (например, СD2). Их рецепторы (ICAM-1, VCАM-1, CD58) вносят вклад в стимуляцию АПК. Благодаря избыточности молекул, поставляющих активационные сигналы, выключение каждой из них вследствие мутаций обычно не приводит к видимым последствиям. Тем не менее нокаут гена CD28 проявляется дефектом Т-хелперов (но не цитотоксических Т-клеток), подавлением образования цитокинов, особенно IL-2 и продуктов ^2-клеток, участвующих в реализации гуморального иммунного ответа. При нокауте гена CD40 подавлен ответ Т-клеток на белковые антигены, причем в большей степени страдает развитие TM-клеток, отвечающих за клеточный иммунный ответ, и выработка ими IFNy. Известна мутация гена CD40L, локализованного в Х-хромосоме, которая в гомозиготном состоянии служит причиной развития первичного иммунодефицитного заболевания — гипер-IgM-синдрома (см. раздел 4.7.1.5).
Источник: Ярилин.А.А , «Иммунология » 2010
А так же в разделе «Костимуляция »
- Презентация антигена
- Миграция клеток, участвующих в презентации антигена
- Иммунный синапс
- Активация Т-лимфоцитов
- Пролиферативная экспансия клонов Т-хелперов
- Дифференцировка Т-хелперов
- Th17 и другие адаптивные субпопуляции Т-клеток
- ИММУННЫЙ ОТВЕТ
- Клеточный иммунный ответ
- Воспалительный Т-клеточный иммунный ответ
- Гуморальный иммунный ответ
- Активация В-лимфоцитов. Роль Т-клеток и цитокинов
- Дифференцировка и селекция В-клеток в зародышевых центрах
- Гибридомы и моноклональные антитела. Генно-инженерные антитела
- Иммунологическая память и вторичный иммунный ответ
- В-клетки памяти
- Т-клетки памяти
- Вторичный иммунный ответ
- Неклассические проявления иммунных реакций
- 3.6.4.3. Проявления активности у5Т- и CD8aa+ Т-клеток
- Иммунологические функции NKT-клеток
- Иммунные процессы в слизистых оболочках (мукозальный иммунный ответ)
- Афферентное и центральное звенья мукозального иммунного ответа
- Роль миграции клеток в мукозальном иммунитете
- Эффекторные механизмы мукозального иммунитета
- Контроль и регуляция иммунного ответа
- Генетический контроль иммунного ответа
- Эндокринный и нервный контроль иммунного ответа
- Регуляция иммунного ответа
- Регуляторные Т-клетки
- РЕЗЮМЕ