Th17 и другие адаптивные субпопуляции Т-клеток
В ходе иммунного ответа CD4+ Т-клетки дифференцируются в нескольких направлениях, в результате чего помимо Th1- и Th2-клеток возникают другие адаптивные субпопуляции Т-хелперов, а также регуляторных Т-лимфоцитов. Вероятно, по времени эти процессы несколько отстают от развития Th1- и ^2-клеток.
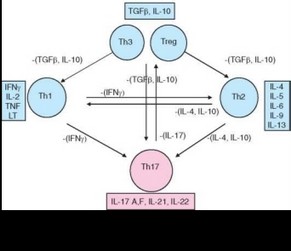
Субпопуляция ThH-лимфоцитов, названных так по их ключевому цитокину IL-17, дифференцируется из активированных CD4+ клеток независимо от Th1- и Th2-лимфоцитов (см. табл. 3.25). Их развитие направляют другие цитокины — IL-6, TGFp, IL-23. Цитокины, секретируемые Th1- и ^2-клетками, подавляют равитие ThH-лимфоцитов. Основным дифференцировочным (транскрипционным) фактором, ответственным за
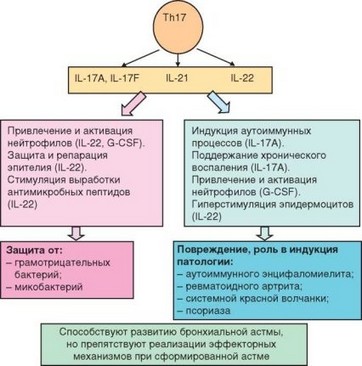
Рис. 3.102. ТЫ7-лимфоциты: механизмы действия, биологические эффекты и значение
развитие Th17-клеток у мыши, служит RORyt (у человека — ROR-C). Эти клетки секретируют IL-21, IL-22 и 2 цитокина семейства IL-17 — IL-17A и IL-17F (рис. 3.102).
Подобно другим типам хелперных Т-лимфоцитов ThH-клетки могут участвовать как в иммунной защите от патогенов, так и в формировании иммунопатологии. Обе этих роли опосредованы преимущественно цитокинами, секретируемыми ThH-клетками, особенно IL-17A и IL-22. Так, эти клетки способны привлекать и активировать нейтрофилы. Это свойство ThH-клеток обусловлено IL-22: он усиливает выработку G-CSF, стимулирующего образование нейтрофилов. Мобилизуя нейтрофилы, ThH-клетки участвуют в защите от грамотрицательных бактерий и в то же время могут способствовать повреждению тканей при хроническом воспалении. Для участия в аутоиммунных процессах необходима присущая ThH-клеткам провоспалительная активность, проявляемая в поддержании хронического, но не острого воспаления, развитие которого ThH-клетки скорее подавляют.
По-видимому, три описанных выше типа адаптивных субпопуляций не исчерпывают всего разнообразия Т-хелперов. Например, в качестве само-
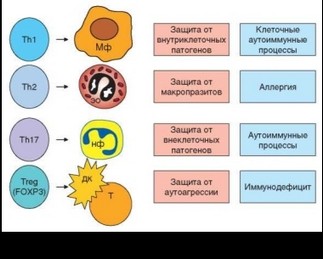
стоятельной субпопуляции некоторые исследователи рассматривают фолликулярные Т-хелперы гуморального иммунного ответа (CD4+ T^-клетки). Сообщалось о CD4+ Т-клетках, специализированных на выработке IL-9 (Th9), а также IL-20 (Th20). Не вызывает сомнений наличие нескольких субпопуляций CD4+ лимфоцитов, выполняющих функции регуляторных Т-клеток, которые будут рассмотрены в соответствующем разделе (см. 3.6.6.3). Таким образом, в настоящее время можно с определенностью говорить о существовании по меньшей мере четырех адаптивных субпопуляций CD4+ Т-лимфоцитов (рис. 3.103, 3.104).
- Цитокины, контролирующие и опосредующие адаптивные реакции лимфоцитов
В главе, посвященной цитокинам (см. раздел 2.5.5), были рассмотрены гуморальные факторы, участвующие в обеспечении реакций врожденного иммунитета, — провоспалительные цитокины, вырабатываемые преимущественно миелоидными клетками. В реакциях адаптивного иммунитета используются цитокины другой группы, которые вырабатываются главным образом лимфоидными клетками (т.е. лимфокины). К ним относят основной ростовой фактор Т-лимфоцитов IL-2, обеспечивающий пролиферативную экспансию клонов Т-клеток, а также цитокины, участвующие в индукции и выполнении функций различных субпопуляций Т-хелперов.
IL-2. Это первый открытый лимфокин, т.е. цитокин, продуцируемый лимфоцитами. Он описан в 1976 г. как фактор роста Т-клеток. В 1983 г. клонирован его ген, содержащий 4 экзона. IL-2 является мономерным гликопротеином с молекулярной массой 15 кДа, содержащим одну дисульфидную связь и один сайт гликозилирования (в гликозилированной форме его молекулярная масса составляет 17—22 кДа).
Основные продуценты IL-2 — CD4+ Т-лимфоциты. Они составляют до 90% клеток-продуцентов этого цитокина. IL-2 образуют также цитотоксические CD8+ T-лимфоциты и некоторые другие клетки. Наиболее активно CD4+ Т-клетки продуцируют IL-2 на начальном этапе после активации (стадия Th0) до дифференцировки на субпопуляции. Из субпопуляций CD4+ Т-клеток (см. выше) его вырабатывают только ТЫ-клетки, но интенсивность секреции IL-2 этими клетками в ходе иммунного ответа постепенно ослабевает. Описано явление супериндукции IL-2 — усиление его выработки ингибиторами белкового синтеза, что объясняется снятием влияния ингибирующих белковых факторов и наличием стабильной матричной РНК. Аналогичное действие оказывает ионизирующая радиация (10—15 Гр).
Субъединичная структура рецептора для IL-2 уже была рассмотрена при иллюстрации зависимости сродства к цитокину от состава рецептора (см. раздел 2.5.5.2). Высокоаффинный рецептор для IL-2 (Kd=10-11 М) содержит 3 полипептидные цепи, из которых 2 — в и у [общая для ряда цитокинов у(с)-цепь] — экспрессируются на Т- и NK-клетках конститутивно (т.е. независимо от активации), тогда как экспрессия a-цепи индуцируется активацией. Все полипептидные цепи рецептора являются трансмембранными белками I типа. При этом за связывание с IL-2 отвечают в- и a-цепи, а за передачу сигнала — у- и в-цепи. И единичная a-цепь, и комбинация с в- и у-цепями обеспечивают связывание цитокина, но в силу низкого сродства рецептора к IL-2 формирующийся комплекс не поглощается клеткой и не вызывает последующей генерации сигнала. Поглощение комплекса важно для разрушения выполнившей свою функцию молекулы цитокина, что препятствует их накоплению, способному привести к нежелательным последствиям. Только тримерный рецептор состава аву имеет необходимый уровень аффинности рецептора к IL-2, что обеспечивает интернализацию комплекса и передачу активационного сигнала. В NK-клетках a-цепь, как правило, не экспрессируется и функциональным является рецептор состава Ру.
Передача сигнала от рецептора для IL-2 происходит по нескольким путям, среди которых основной и наиболее специфичный — путь, начинающийся при активации киназ семейства Jak, связанных с в- и у-цепями рецептора. С в-цепью связана киназа Jak1, c у-цепью — Jak3. Конформационные изменения этих цепей вызывают активацию Jak-киназ, реализующих свое действие через фосфорилирование факторов STAT3 и STAT5. Эти факторы димеризуются, мигрируют в ядро и взаимодействуют с участками промоторов различных генов, контролирующих клеточный цикл, выступая в качестве дифференцировочных факторов. Для связывания с киназами предназначены специальные участки в цитоплазматической части цепей рецептора, обогащенные отрицательно заряженными (содержащими карбоксилы) аминокислотными остатками. Под влиянием Jak-киназ происходит фосфорилирование этих остатков. Это обеспечивает возможность взаимодействия цитоплазматических участков цепей с киназой Lck, участвующей в запуске другого сигнального пути, в который вовлечена Ras/Raf-зависимая ветвь MAP-каскада. Это приводит к формированию дополнительных дифференцировочных факторов, участвующих во включении генов, необходимых для продвижения клеток по клеточному циклу. При участии Lck происходит также активация липидной киназы PI3K, активирующей белок Vav — основной фактор, ответственный за включение цепи реакций, обеспечивающих необходимую для прохождения митоза перестройку цитоскелета.
IL-2 вызывает два основных физиологических эффекта — индуцируют антигензависимую пролиферацию всех разновидностей Т-клеток и способствуют дифференцировке некоторых функциональных субпопуляций лимфоцитов — цитотоксических лимфоцитов, регуляторных Т-клеток. Спектр мишеней этого фактора довольно узок. К ним относят Т-лимфоциты и естественные киллеры, а также В-лимфоциты. IL-2 применяют в качестве главного фактора для поддержания длительной пролиферации Т-клеток с последующим их клонированием и получением постоянных культур клонов Т-лимфоцитов. Особенно чувствительны к действию IL-2 цитотоксические CD8+ Т-лимфоциты, для дифференцировки которых он необходим. IL-2 усиливает цитотоксическую эффективность NK-клеток, существенно расширяя спектр мишеней, на которые действуют формирующиеся под его влиянием клетки (LAK-клетки — см. раздел 4.1.2.3). IL-2 выступает в качестве кофактора пролиферации активированных В-лимфоцитов, способствует повышению функциональной активности моноцитов, экспрессирующих рецептор для IL-2 состава ву. Наконец, IL-2 влияет на гемопоэз, усиливая образование тромбоцитов и эозинофилов и ослабляя миело- и эритропоэз, а также способствуя формированию экстрамедуллярных очагов гемопоэза.
Роль IL-2 в качестве ростового фактора активированных Т-клеток, еще недавно считавшаяся главной функцией IL-2, заменима, поскольку аналогичный эффект могут дать IL-7, IL-4, а также комбинации провоспалительных цитокинов. В то же время IL-2 служит незаменимым фактором индукции дифференцировки естественных регуляторных Т-лимфоцитов и главным фактором роста этих клеток. При нокауте гена IL2 проявляются последствия повреждения именно этой функции данного цитокина. Следствием выключения IL-2 является развитие доброкачественных лимфопролиферативных процессов, захватывающих преимущественно кишечник, а также аутоиммунной полиэндокринопатии и ряда других аутоиммунных нарушений. По не вполне понятным причинам ростсти- мулирующее действие IL-2 в отношении Трег четко проявляется in vivo и с трудом воспроизводится в клеточной культуре. Изменение предоставлений о ключевых функциях IL-2 существенно ограничило использование этого цитокина как средства иммунотерапии (см. раздел 4.8.3.2).
Цитокины IL-12 и IFNy рассмотрены в главе 2 в контексте врожденного иммунитета.
IL-4. Описан в 1981 г. как фактор, стимулирующий активированные В-лимфоциты. Гликопротеин с молекулярной массой 15,5 кДа (гликозили- рованная форма — 19—22 кДа), содержит 1 функционально важную дисульфидную связь.
Основные продуценты IL-4 — ТИ2-клетки. Как отмечалось выше, его вырабатывают также тучные клетки, базофилы, эозинофилы, NK- и NKT-клетки, а также дендритные клетки (DC-2) — преимущественно спонтанно. IL-1, IL-2, а также глюкокортикоиды усиливают выработку IL-4.
Рецепторы IL-4 содержат 2 цепи — а и у(с). Полипептидные цепи рецептора относят к семейству цитокиновых (гемопоэтиновых) рецепторов. Основные мишени IL-4 — В-лимфоциты. На одной покоящейся В-клетке содержится 200—300 высокоаффинных рецепторов; их число значительно возрастает после активации.
IL-4 служит основным ростовым фактором для В-лимфоцитов. Он может вызвать их пролиферацию без дополнительных стимулов. IL-4 способен вызвать переключение С-генов IGH на изотипы у1 и е, что усиливает выработку антител классов IgG1 (только у мышей) и IgE. IL-4 вызывает усиление экспрессии на В-клетках низкоаффинного рецептора FceRII (CD23), растворимая форма которого стимулирует синтез IgE. Усиливающее действие на синтез IgE и способность поддерживать пролиферацию тучных клеток (преимущественно серозных) обусловливает участие IL-4 в патогенезе аллергических заболеваний.
Для Т-лимфоцитов IL-4 служит «запасным» ростовым фактором: он может замещать ростовое действие IL-2. К этому действию IL-4 чувствительны CD8+ Т-лимфоциты и ТИ2-клетки, для которых он служит основным фактором роста. Кроме того, IL-4 — единственный цитокин, способный индуцировать дифференцировку ТИ2-клеток. Действуя на CD8+ Т-клетки, он индуцирует развитие цитотоксических Т-лимфоцитов, секретирующих IL-4 и другие ТИ2-цитокины.
IL-4 служит функциональным антагонистом IFNy, как при непосредственном действии на клетки (он подавляет выработку IFNy и ослабляет некоторые его эффекты), так и при индукции образования ТИ2-клеток. Однако в некоторых случаях (например, при индукции и усилении экспрессии на клетках молекул MHC-II) эти цитокины проявляют синергизм. Сочетание антагонизма и синергизма выявляют и в функциональных взаимоотношениях IL-4 с IL-2.
Подавляя активность макрофагов и синтез ими IL-1, TNFa, IL-6 и других провоспалительных цитокинов, IL-4 выступает в роли противовоспалительного цитокина. В то же время он усиливает цитотоксическую активность макрофагов и миграцию нейтрофилов в очаг воспаления. Выступая в качестве основного фактора аллергических процессов, он способствует
развитию аллергического воспаления. Кроме того, IL-4 стимулирует гемопоэз, в частности выработку колониестимулирующих факторов. Описаны его противоопухолевые эффекты.
Таким образом, IL-4 возглавляет список ^2-цитокинов и факторов, индуцирующих неклассический тип воспаления — аллергическое воспаление. Он служит также главным ростовым цитокином В-клеток и фактором, ответственным за переключение изотипов, тем самым способствуя развитию гуморального иммунного ответа.
IL-5. Описан как дифференцировочный фактор В-лимфоцитов. Он представляет собой гомодимер с молекулярной массой 45—60 кДа. IL-5 имеет 3 участка гликозилирования. При массе каждой цепи в 22,5 кДа, только 12,5 кДа приходится на белковую часть.
Главный источник IL-5 — ^2-клетки. Его секретируют также тучные клетки и эозинофилы. Основные мишени IL-5 — эозинофилы и активированные В-клетки (активация способствует повышению числа высокоаффинных рецепторов на клетках). Способность IL-5 поддерживать пролиферацию В-лимфоцитов реализуется на более поздних стадиях развития, чем аналогичный эффект IL-4. Это действие в значительной степени косвенное — IL-5 усиливает экспрессию рецепторов для IL-2. IL-5 стимулирует дифференцировку В-лимфоцитов в плазматические клетки. Не влияя на переключение изотипов, этот цитокин повышает число продуцентов IgM, а также, действуя на посттранскрипционном уровне, усиливает выработку IgA. Тем самым IL-5 способствует защите слизистых оболочек, в которых IgA составляет основной изотип антител.
Однако главное проявление биологического действия IL-5 — его влияние на эозинофилы: он служит фактором выживаемости, роста, дифференци- ровки и хемотаксиса этих клеток. Выделяемый тучными и ^2-клетками в очаге аллергической реакции, IL-5 представляет главный фактор, ответственный за развитие отложенной фазы аллергической реакции немедленного типа, центральным событием которой является миграция в очаг поражения эозинофилов и некоторых других форм лейкоцитов.
Таким образом, IL-5, являясь цитокином с относительно узким спектром биологической активности, влияет на эозинофилы и В-клетки, продвинутые по пути дифференцировки в плазматические клетки.
IL-13. IL-13 относят к группе ^2-цитокинов. Его молекулярная масса составляет всего 10 кДа. Ген этого цитокина расположен на 5-й хромосоме в составе кластера ^2-цитокинов (наряду с IL-4, IL-5, IL-3, IL-9, IL-13, GM-CSF). Вероятно, этот ген является продуктом дупликации гена IL4. Вначале IL-13 считали «заместителем» IL-4, поскольку он способен воспроизводить многие его эффекты. Основой для этого предположения, наряду со структурной гомологией, послужило сходство строения рецепторов этих цитокинов, имеющих общую a-цепь (IL-R4a), но вместо у(с)-цепи (входит в состав рецептора IL-4) рецептор для IL-13 содержит собственную a-цепь — IL-13Ra.
IL-13 вырабатывают ^2-клетки, эозинофилы и тучные клетки. Спектр его эффектов весьма широк — мишенями IL-13 служат не только В-лимфо- циты и эозинофилы (что обычно для ^2-цитокинов), но и гладкомышечные клетки, продуцирующие слизь бокаловидные и другие эпителиальные клетки слизистых оболочек. Будучи способным воспроизводить большинство эффектов IL-4 при аллергии немедленного типа, IL-13 может выполнять при этих реакциях также более специальные функции. Так, он отвечает за усиление продукции слизи, спазм гладкой мускулатуры бронхов и, наконец, за изменение строения стенки бронхов, обозначаемое как ремоделирование (см. раздел 4.5.1.5.). В результате IL-13 рассматривают как наиболее важный и универсальный по спектру действия эффектор аллергических реакций немедленного типа.
Источник: Ярилин.А.А , «Иммунология » 2010
А так же в разделе «Th17 и другие адаптивные субпопуляции Т-клеток »
- Презентация антигена
- Миграция клеток, участвующих в презентации антигена
- Иммунный синапс
- Костимуляция
- Активация Т-лимфоцитов
- Пролиферативная экспансия клонов Т-хелперов
- Дифференцировка Т-хелперов
- ИММУННЫЙ ОТВЕТ
- Клеточный иммунный ответ
- Воспалительный Т-клеточный иммунный ответ
- Гуморальный иммунный ответ
- Активация В-лимфоцитов. Роль Т-клеток и цитокинов
- Дифференцировка и селекция В-клеток в зародышевых центрах
- Гибридомы и моноклональные антитела. Генно-инженерные антитела
- Иммунологическая память и вторичный иммунный ответ
- В-клетки памяти
- Т-клетки памяти
- Вторичный иммунный ответ
- Неклассические проявления иммунных реакций
- 3.6.4.3. Проявления активности у5Т- и CD8aa+ Т-клеток
- Иммунологические функции NKT-клеток
- Иммунные процессы в слизистых оболочках (мукозальный иммунный ответ)
- Афферентное и центральное звенья мукозального иммунного ответа
- Роль миграции клеток в мукозальном иммунитете
- Эффекторные механизмы мукозального иммунитета
- Контроль и регуляция иммунного ответа
- Генетический контроль иммунного ответа
- Эндокринный и нервный контроль иммунного ответа
- Регуляция иммунного ответа
- Регуляторные Т-клетки
- РЕЗЮМЕ