Иммунный синапс
Второе из сформулированных выше препятствий для осуществления презентации — дефицит молекул MHC, несущих «нужные» пептиды, — решается благодаря формированию особой структуры, необходимой для успешной презентации антигена — иммунного синапса или супрамолекулярного активационного кластера (SMAC — Supramolecular activation cluster). Иммунный синапс — структурированная зона контакта между клетками, участвующими в реализации той или иной формы иммунологического распознавания и связанной с ним передаче сигнала. Иммунный синапс формируется с участием зрелой дендритной клетки и CD4+ Т-лимфоцита для презентации антигена и представляет наиболее многокомпонентную форму этого процесса. С участием иммунного синапса реализуются 3 основных условия эффективной презентации:
- устраняются стерические помехи для взаимодействия клеток;
- обеспечивается мобилизация молекул адгезии, необходимых для формирования контакта между клетками и его стабилизации;
- оптимизируется передача активирующего сигнала.
Выделяют 3 стадии формирования иммунного синапса — поляризацию клеток, установление зоны первичного контакта между клетками и образование зрелого иммунного синапса, способного обеспечить передачу сигнала. Поляризация клеток происходит в процессе их сближения, направляемого хемокинами. Как уже упоминалось, дендритные клетки привлекают Т-хелперы, выделяя СС-хемокины CCL19 и CCL21, распознаваемые Т-хел- перами при помощи рецептора CCR7. Это обеспечивает поляризацию клеток, направленное движение Т-хелперов к дендритным клеткам и служит условием установления контакта между ними.
Поляризация клеток заключается в ориентации мембранных и внутриклеточных компонентов таким образом, чтобы облегчить не только установление контакта, но и последующий обмен сигналами, необходимыми для их взаимной активации. Участок клетки, расположенный в направлении ее движения, называют лидирующим, противоположный конец — хвостовым. В поляризации клеток участвуют актинсодержащие компоненты цитоскелета, активируемые сигналами от хемокиновых рецепторов (см. раздел 2.3.2.2). При перестройке цитоскелета происходит переориентация клеточного центра, организующего микротрубочки, локальная полимеризация актина и латеральное перемещение белков, обеспечивающее их накопление в зоне контакта клеток. Полимеризация актина происходит вследствие реализации цепи событий, запускаемых хемокинами и активацией G-белков, связанных с их рецепторами. В этом процессе участвуют фактор Vav и белок WASP, образующий комплекс с белком ARP2/3. В основе поляризации и направленного движения Т-хелперов и дендритных клеток лежат те же механизмы, что и при хемотаксисе фагоцитов.
Для формирования первичного контакта между клетками необходима остановка движения Т-лимфоцитов, зависимая от сигналов, исходящих от TCR и корецептора CD4. Остановка происходит уже через 30 с после начала формирования контакта. Другое условие сближения клеток состоит в особом перераспределении мембранных молекул, которое достигается как в результате поляризации клеток, так и под влиянием событий, связанных с самим контактом. Прежде всего происходит сортировка молекул по размеру. Крупные молекулы, такие как CD43 и CD45, мешают сближению клеток из-за своих размеров: протяженность (длина) этих молекул составляет около 40 нм, тогда как для передачи сигнала клетки должны сблизиться на расстояние 5—15 нм. Кроме того, с этими молекулами связана большая
часть отрицательного заряда клеток, определяющего их взаимное отталкивание. Благодаря перераспределению молекул гликопротеин CD43 сосредотачивается в хвостовом отделе клетки. Молекула CD45 также выводится из лидирующего участка, но лишь временно, поскольку она участвует в передаче сигнала.
В лидирующем участке сосредоточиваются молекулы адгезии: на Т-хел- пере — в2-интегрин LFA-1, на дендритной клетке — его рецептор ICAM-1. Формирование комплексов между этими молекулами составляет основу первичного контакта между клетками. Дополнительный вклад в этот процесс вносит взаимодействие молекул CD2 Т-хелпера и CD58 (LFA-3) дендритной клетки. Зона адгезивного взаимодействия LFA-1—ICAM-1 окружена молекулами, которым предстоит сыграть основную роль в презентации антигена: на дендритной клетке — MHC-II, содержащими распознаваемый пептид, а на Т-хелпере — apTCR (в начале взаимодействия он находится в хвостовой части клетки, но быстро перемещается в зону контакта).
Цель формирования зрелого синапса состоит в том, что молекулы центрального и периферического участков зоны контакта меняются местами: на Т-хелпере молекулы TCR перемещаются в центр, вытесняя молекулы LFA-1 на периферию синапса, а на дендритной клетке аналогичным образом комплексы MHC-II—пептид меняются местами с молекулами ICAM-1. При моделировании иммунного синапса с использованием вместо АПК искусственных мембран, содержащих молекулы, меченные флуоресцентными красителями, удается визуализировать структуру иммунного синапса, поскольку его центральная и периферическая зоны окрашиваются разными флуорохромами. Наблюдаемую при этом характерную структуру обозначают как «бычий глаз» (рис. 3.86).
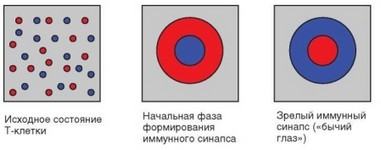
Рис. 3.86. Схема формирования иммунологического синапса. При распознавании Т-клеточным рецептором комплекса молекула MHC—пептид происходит перераспределение мембранных молекул: диффузное распределение сменяется иммунным синапсом, центр которого вначале занят молекулами адгезии, а затем — специфическими комплексами ТСК-(МНС-пептид). Красным цветом обозначена экспрессия рецепторного комплекса TCR—CD3, синим — интегрина LFA-1
Помимо указанных перемещений созревание синапса подразумевает привлечение в центральную его часть полного набора молекул, участвующих в восприятии и передаче антигенного сигнала — корецепторов, костимулирующих молекул, связанных с мембранами киназ и адапторных белков. Важная роль в перераспределении молекул в зоне формирования иммунного синапса принадлежит рафтам. Рафтами (от англ. raft — плот) называют нерастворимые в детергентах субъединицы (микродомены) мембраны, обогащенные холестерином и сфинголипидами. Только некоторые мембранные белки включены в состав рафтов и перемещаются по мембране вместе с ними. К этим белкам на поверхности Т-хелперов принадлежат корецепторы CD4 и CD8, тирозинкиназа Lck (ассоциирована с CD4 и CD8), костимулирующие молекулы (включая CD28), адапаторный белок LAT, тирозинкиназа ZAP-70, PLCy, PI3K, а также тирозинфосфатаза CD45, возвращающаяся в зону контакта после предварительного удаления из нее. В дендритных клетках в состав рафтов входят молекулы MHC-II и костимулирующие молекулы CD80/86.
В то же время молекулярный комплекс, который является ключевым в рассматриваемом процессе — TCR — в покоящихся Т-клетках с рафтами не связан. Его вовлечение в иммунный синапс обусловлено формированием нековалентной связи между TCR и CD4, вследствие чего CD4 как бы втягивает рецептор в рафт и тем самым обеспечивает его включение в состав иммунного синапса. В состав рафтов входят также изоформа 0 протеинкиназы С, вовлекаемая в рафты благодаря установлению связей с другими звеньями сигнального пути. Накопление этой молекулы в иммунном синапсе рассматривают как проявление его «зрелости».
В конечном счете на малых участках поверхности Т-хелпера и дендритной клетки, обращенных друг к другу, концентрируются практически все комплексы MHC-II—пептид, а также значительная часть молекул TCR, CD4, костимулирующих и сигнальных молекул, необходимых для активации Т-хелпера (рис. 3.87). Признаки зрелости синапса можно зарегистрировать уже через 5 мин, но максимальная плотность молекул, участвующих в презентации антигена, достигается через 10—20 мин.
Сродство TCR к комплексу MHC-II—пептид очень невелико: Kd взаимодействия этих молекул составляет 10-4—10-6 М. Вовлечение в распознавание
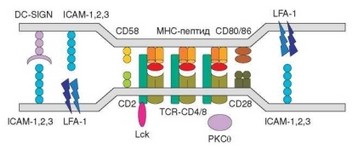
Рис. 3.87. Структура иммунного синапса: детализированное изображение зрелого иммунного синапса
корецептора CD4 повышает эту величину на 2 порядка (т.е. до 10"6—10"8 М), что все равно не делает ее достаточно высокой. Как уже упоминалось, для индукции активации необходимо вовлечение 200—500 пар молекул, в то время как число молекул MHC, несущих «нужный» (т.е. распознаваемый данным TCR) пептид, составляет менее 100. Благодаря концентрации молекул MHC, несущих специфических пептид, в зоне синапса на площади около 0,45 мкм2 (при объеме 10-18 л), плотность комплексов TCR—(MHC— пептид) составляет 100—350 на 1 мкм2. Аккумуляция комплексов в иммунном синапсе тем больше, чем выше сродство пептида к TCR. Высокая плотность комплексов способствует повышению эффективности распознавания. Другая важная характеристика синапса — время полужизни комплексов TCR—(MHC—пептид), зависящее от сродства пептида к молекуле МНС-II. Если пептид, связанный с МНС, является сильным агонистом, время полужизни комплекса составляет 10 с и больше, тогда как аналогичное время для комплексов, содержащих слабый агонист, составляет меньше 10 с.
Однако наиболее важно для повышения эффективности презентации то, что каждый комплекс MHC-II—пептид (т.е. лимитирующий компонент презентации) может распознаваться молекулами TCR повторно. После довольно продолжительного взаимодействия TCR и комплекса MHC—пептид, необходимого для завершения фосфорилирования Z-цепи рецептора и передачи сигнала в клетку, молекула TCR интернализуется (поглощается клеткой), а на ее место поступает другая молекула TCR, и все события повторяются. Показана возможность взаимодействия одного комплекса MHC-II—пептид с 200 молекулами TCR за 1 ч. Одна из функций иммунного синапса состоит как раз в обеспечении серийного распознавания комплексов MHC—пептид и продолжительной (до 20 ч при активации наивных Т-клеток и только 1 ч — при активации Т-клеток памяти) передачи сигнала, необходимой для активации Т-хелперов.
Так, запуск комплекса механизмов, реализуемых на уровне целого организма (улавливание клонов Т-клеток в региональном лимфоузле) и на уровне клетки (перераспределение мембранных молекул взаимодействующих клеток с формированием иммунного синапса), позволяет преодолеть трудности, возникающие на пути эффективной презентации антигена Т-клеткам.
Описанный выше процесс презентации антигена дендритными клетками Т-хелперам можно рассматривать как прототип разнообразных аналогичных процессов, реализуемых на разных этапах иммунного ответа. Те же закономерности лежат в основе презентации антигена дендритными клетками цитотоксическим Т-лимфоцитам с той разницей, что презенти- руемый антигенный пептид образует комплекс не с MHC-II, а с MHC-I и в качестве корецептора выступает не CD4, а CD8. Аналогичным образом осуществляется презентация антигенных пептидов Т-клеткам макрофагами и В-лимфоцитами в ходе иммунного ответа. Но в этом взаимодействии принимают участие не наивные, а преактивированные Т-клетки, что несколько упрощает процедуру формирования иммунного синапса и генерации активационных сигналов. Наконец, определенные события презентации реализуются при взаимодействии цитотоксических лимфоцитов с клетками-мишенями. Тем не менее, в наиболее полной, классической форме межклеточное взаимодействие рассматриваемого типа проявляется именно при презентации антигена дендритными клетками наивным CD4+ Т-лим- фоцитам.
Источник: Ярилин.А.А , «Иммунология » 2010
А так же в разделе «Иммунный синапс »
- Презентация антигена
- Миграция клеток, участвующих в презентации антигена
- Костимуляция
- Активация Т-лимфоцитов
- Пролиферативная экспансия клонов Т-хелперов
- Дифференцировка Т-хелперов
- Th17 и другие адаптивные субпопуляции Т-клеток
- ИММУННЫЙ ОТВЕТ
- Клеточный иммунный ответ
- Воспалительный Т-клеточный иммунный ответ
- Гуморальный иммунный ответ
- Активация В-лимфоцитов. Роль Т-клеток и цитокинов
- Дифференцировка и селекция В-клеток в зародышевых центрах
- Гибридомы и моноклональные антитела. Генно-инженерные антитела
- Иммунологическая память и вторичный иммунный ответ
- В-клетки памяти
- Т-клетки памяти
- Вторичный иммунный ответ
- Неклассические проявления иммунных реакций
- 3.6.4.3. Проявления активности у5Т- и CD8aa+ Т-клеток
- Иммунологические функции NKT-клеток
- Иммунные процессы в слизистых оболочках (мукозальный иммунный ответ)
- Афферентное и центральное звенья мукозального иммунного ответа
- Роль миграции клеток в мукозальном иммунитете
- Эффекторные механизмы мукозального иммунитета
- Контроль и регуляция иммунного ответа
- Генетический контроль иммунного ответа
- Эндокринный и нервный контроль иммунного ответа
- Регуляция иммунного ответа
- Регуляторные Т-клетки
- РЕЗЮМЕ