Дифференцировка Т-хелперов
Под влиянием активации в результате изменений структуры хроматина гены цитокинов становятся доступными для регулирующих сигналов, под влиянием которых выявляют минимальный уровень экспрессии многих из этих генов. На этом этапе CD4+ Т-клетки обозначаются как Th0-клетки. Помимо IL-2, обеспечивающего пролиферативную экспансию, в этих клетках слабо экспрессируются IL-3, IL-4, IL-5, IL-6, IL-9, IL-10, IL-13, GM-CSF, IFNy, TNFa и некотрые другие факторы, что на данном этапе, по-видимому, не играет существенной функциональной роли. Но уже в процессе деления CD4+ Т-клеток запускается процесс их дифференцировки на субпопуляции. Такие субпопуляции называют адаптивными, поскольку они образуются в ходе адаптивного иммунного ответа на антигены, в отличие от естественных субпопуляций (см. раздел 3.3.2.1), формирующихся в ходе антигеннеза- висимой дифференцировки Т-клеток (табл. 3.25). Ранее других образуются 2 субпопуляции Т-хелперов — Th1 и Th2 (Th — от T-helper).
Дихотомию Т-хелперного звена системы адаптивного иммунитета впервые описали в 1986 г. Т. Мосманн (T.R. Mosmann) и соавт. Они обнаружили, что панель Т-хелперных клонов мышей по способности секретировать цитокины можно разделить на 2 группы, и обозначили их как Th1 и Th2.
Таблица 3.25. Адаптивные субпопуляции Т-хелперов
Первые секретировали IFNy, IL-2, IL-3, GM-CSF, вторые — IL-4, IL-5 и IL-3. Позже дифференцировка Т-хелперов на 2 субкласса была показана in vivo при иммунном ответе и была продемонстрирована функциональная значимость Th1/Th2-дихотомии. Этот важный вывод был сделан при анализе природы различий в чувствительности к Leishmania major мышей линий BALВ/c (чувствительная линия) и C57BL/6 (устойчивая линия). Оказалось, что у мышей C57BL/6 в ответ на инфицирование лейшманиями формируются преимущественно Th1-, а у мышей BALB/c — ^2-клетки. Различие спектра гуморальных продуктов, прежде всего ключевых цитокинов Th1- и ^2-клеток (соответственно IFNy и IL-4) определяет основную направленность иммунного ответа в направлении клеточной или гуморальной защиты. Клеточный иммунный ответ обусловливает эффективную защиту от внутриклеточных патогенов (к которым относят лейшмании), а гуморальный ответ — защиту от внеклеточных патогенов. Только соответствие направления дифференцировки Т-хелперов (а следовательно, типа ответа) природе патогена делает эту защиту эффективной. Поэтому мыши линии C57BL/6, формирующие Th1-зависимый клеточный ответ, эффективно защищаются от лейшманиазной инфекции, а мыши линии BALB/c, реагирующие развитием ^2-зависимого гуморального ответа, оказываются беззащитными и гибнут.
Открытие адаптивных субпопуляций Т-хелперов создало основу для формирования представлений о различии клеточных механизмов иммунной защиты, в зависимости от локализации патогена (внутри- или внеклеточная). Это сильно повлияло также на представления о синтезе цитокинов (двойственность набора цитокиновых продуктов показана не только для Т-хелперов, но и для ряда других клеток иммунной системы) и послужило толчком для изучения путей дифференцировки Т-хелперов. Наконец, это открытие оказало значительное влияние на понимание природы таких форм иммунопатологии, как аллергия и аутоиммунные процессы, в развитии которых важную роль играет дисбаланс субпопуляций Т-хелперов. При всей важности этой концепции необходимо сознавать, что она в значительной степени схематизирует иммунологические процессы, что особенно заметно проявляется при рассмотрении иммунных процессов, происходящих в организме человека.
При дифференцировке Т-хелперов происходит супрессия одних и усиление экспрессии других цитокиновых генов, прежде слабо экспрессированных в ThO-клетках. ТЫ-клетки продуцируют IFNy, IL-2, TNFa, TNFp, IL-3, GM-CSF, Th2-клетки - IL-4, IL-13, IL-5, IL-6, IL-9, IL-11, IL-21, IL-25, IL-10, IL-3, GM-CSF. В этих спектрах есть цитокины, общие для двух линий (GM-CSF, IL-3, у человека — IL-10). Другие цитокины более или менее специфичны для субпопуляций. Среди этих цитокинов выделяют ключевые для Th1- и ^2-клеток, поскольку они не просто специфичны для них, но и причастны к выполнению их основных эффекторных функций, индукции дифференцировки клеток этих субпопуляций, самоподдержанию и подавлению развития клеток противоположного типа. Для ТЫ-клеток таким цитокином является IFNy, для ^2-клеток — IL-4 (рис. 3.100).
Факторы и механизмы дифференцировки Th1- и ^2-клеток изучены достаточно детально. Их дифференцировка зависит от действия по меньшей мере двух сигналов. Один из них поступает через TCR и «сообщает» о распознавании антигена в комплексе с молекулой MHC. Второй сигнал поставляют цитокины. Относительная роль этих сигналов до конца не определена. Цитокинам приписывают селективную или инструктивную функции. Согласно селективной точке зрения, направление дифференци- ровки Т-хелперов задается при передаче сигнала через TCR, а цитокины только выбирают ту или иную субпопуляцию, обеспечивая ее факторами выживания и пролиферации. Согласно инструктивной точке зрения, через TCR передается активационный сигнал, тогда как направление дифферен- цировки задают цитокины через соответствующие рецепторы. В настоящее время предпочтение отдают первой точке зрения.
Развитию ТЫ-клеток способствуют крайние (очень высокая и очень низкая) дозы антигена и его высокое сродство к рецептору, а развитию ^2-клеток — промежуточные дозы антигена и более низкое его сродство
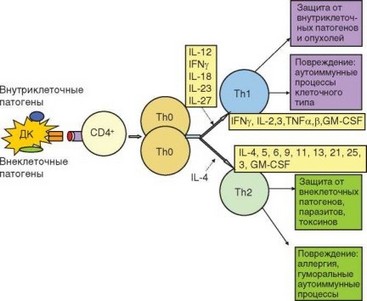
Рис. 3.100. Дифференцировка Т-хелперов типов Th1 и Th2. Указаны пути диффе- ренцировки Т-хелперов двух основных типов, функции образующихся клеток, а также цитокины, направляющие эту дифференцировку и секретируемые самими Т-хелперами
к рецептору. О значении корецептора CD4 в выборе пути дифференциров- ки свидетельствует усиление дифференцировки Т-хелперов в направлении TM-клеток при нокауте гена CD4. С решающей ролью в дифференциров- ке субпопуляций Т-хелперов антигенного сигнала можно связать то, что способность вызывать разные формы иммунного ответа через индукцию ТЫ- или ^2-клеток обусловлена самой природой антигенов. Так, преимущественную дифференцировку TM-хелперов вызывают антигены лейшма- ний, трипаносом, хламидий, микобактерий, бордетелл, бореллий, гелико- бактера, кандид, вируса гриппа, а также бактериальная ДНК, содержащая последовательность CpG, двуспиральная РНК, ЛПС и суперантигены. Избирательными индукторами ^2-хелперов служат антигены шистосом, токсокар, вирусов оспы, кори, а также аллергены и протеазы паразитов. Перечень антигенов-индукторов ТЫ-клеток наводит на мысль, что за TM-ориентацию иммунного ответа отвечают не антигены как таковые, а PAMP, с которыми они связаны. Антигенные эпитопы и детерминанты PAMP могут располагаться на одной молекуле, как например в случае ЛПС. Сигналы от PAMP могут воспринимать рецепторы АПК, прежде всего дендритных, в значительной степени определяющие характер передачи сигнала через антигенраспознающие рецепторы лимфоцитов. В случае антигенов, индуцирующих преимущественно ^2-ответ, молекулярные основы данного предпочтения не установлены. Таким образом, роль антигенов в выборе пути дифференцировки Т-хелперов скорее всего связана с передачей сигнала не через TCR, а через костимулирующие молекулы. Вероятно, влиянием на АПК можно объяснить различное действие адъювантов на диф- ференцировку Т-хелперов. Так, полный адъювант Фрейнда (содержащий микобактерии) способствует развитию Thl-клеток, а алюминиевые квасцы и столбнячный анатоксин — дифференцировке по ^2-пути.
Роль костимуляции, осуществляемой АПК, в выборе пути дифференци- ровки Т-хелперов проявляется при анализе на уровне конкретных костиму- лирующих молекул. Так, преобладающее участие в костимуляции молекул Т-клетки ICOS и OX40, а также молекулы CD86, экспрессированной на дендритной клетке, направляет дифференцировку в сторону ^2-клеток, в то же время костимуляция через CD80 не оказывает существенного влияния на дифференцировку. Предварительная обработка дендритных клеток IFNy или IL-12 придает им способность преимущественно индуцировать ТЫ-дифференцировку, а обработка IL-10 или фактором TSLP — развитие ^2-клеток. Миелоидные дендритные клетки имеют большую способность к индукции дифференцировки Thl-лимфоцитов, а плазмоцитоидные — ^2-клеток. В связи с этим сформировались представления о двух субпопуляциях дендритных клеток — DC1 и DC2, индуцирующих развитие соответственно Th1- и ^2-клеток (см. раздел 2.1.6).
Роль цитокинов в обеспечении дифференцировки Th1- и ^2-клеток изучена более детально. К настоящему времени установлено, что основным индуктором Thl-клеток служит IL-12. Развитию и поддержанию Thl-ответа способствуют также IFNy, IL-18, IL-23 и IL-27. Аналогичную, но второстепенную роль играют TNFa, TGFp, IFNa. Цитокином, определяющим развитие ^2-клеток, служит только IL-4. Роль второстепенных факторов при их развитии играют IL-10, IL-6, IL-2, IL-19 и даже цепь р40 IL-12. Th0-клетки экспрессируют рецепторы для названных цитокинов. Они несут «полные» рецепторы для IL-12 (PiP2), IFNy (aP) и IL-4 (ay), через которые получают сигналы, определяющие дальнейшую судьбу Т-хелпера. Уже после дифференцировки на некоторых клетках структура названных рецепторов становится неполной. Так, ^2-клетки утрачивают р2-цепь рецептора для IL-12. Полный рецептор для IFNy сохраняют только ^2-клетки, тогда как Th1-лимфоциты утрачивают р-цепь.
Влияние IL-12 на дифференцировку Th1-клеток реализуется через рецептор, содержащий 2 полипептидные цепи — Pj и р2. Цитоплазматические участки цепей связаны с тирозинкиназами семейства Jak: Pj-цепь связана с киназой Tyk2, а р2-цепь — с киназой Jak2. Эти киназы фосфорилируют (и, следовательно, активируют) транскрипционные факторы STAT1, STAT3, STAT4 и STAT5. Показано, что наиболее важен для дальнейшей передачи сигнала тандем р2-цепи с киназой Jak2. Именно он отвечает за фосфорилирование транскрипционного фактора STAT4, которому принадлежит ключевая роль в передаче сигнала, приводящего к дифференцировке Thl-клеток. В активации STAT4 задействован также сигнальный путь, реализуемый через MAP-киназу р38, которая фосфорилирует STAT4. Среди эффектов STAT4 следует выделить его способность взаимодействовать с промотором гена, кодирующего IFNy, и запускать его экспрессию.
Другой транскрипционный фактор, играющий ключевую роль в формировании Thl-фенотипа — T-bet (рис. 3.101). Его экспрессия коррелирует с продукцией IFNy. Экспрессия этого фактора зависит от сигналов, поступающих от TCR и IFNyR, но не зависит от сигналов, индуцируемых IL-12. Нокаут гена T-bet приводит к полному подавлению выработки IFNy и нарушению дифференцировки Th1-клеток при активации in vitro и in vivo. Выработка ^2-цитокинов при этом усиливается. Таким образом, IFNy усиливает экспрессию T-bet и в то же время экспрессия IFNy зависит от T-bet. Формируется петля положительной обратной связи и достигается стабилизация фенотипа Th1-клеток. В дифференцировке TM-клеток участвуют еще несколько факторов — Hlx, ERM, IRF-1.
Основные транскрипционные факторы, определяющие дифференци- ровку ^2-клеток, — STAT6 и GATA-3 (последнему принадлежит ведущая роль) (см. рис. 3.101). STAT6 активируется при связывании IL-4 со своим рецептором. Активация GATA-3 зависит как от сигнализации через TCR, так и от активности STAT6. GATA-3 «включает» петлю аутоактивации, которую рассматривают как механизм стабилизации фенотипа ^2-клеток. Совместно с дополнительными факторами, в частности сMaf и NF-AT, экспрессируемыми при участии STAT6, GATA-3 индуцирует экспрессию генов IL4, IL5 и IL13. В опытах с нокаутом гена GATA-3 показана однозначная связь индукции ^2-цитокинов с экспрессией GATA-3. Кроме того, GATA-3 супрессирует развитие Th1-клеток, подавляя экспрессию генов STAT4 и IL-12RP2, что затрудняет передифференцировку ^2-лимфоцитов в Th1. GATA-3 участвует в поддержании пролиферации ^2-клеток. Таким
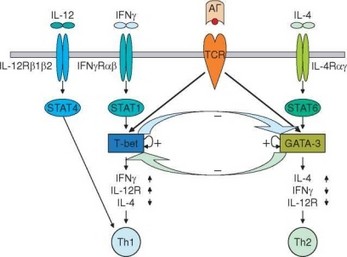
Рис. 3.101. Вклад цитокинов и дифференцировочных факторов в образование Th1- и 1Ь2-лимфоцитов. Отражены основные внеклеточные и внутриклеточные факторы, определяющие дифференцировку Т-хелперов и их взаимное подавление
образом, фактор GATA-3 важен не только для дифференцировки ТИ2-кле- ток, но и для стабилизации их фенотипа, а также подавления программы дифференцировки TW-лимфоцитов.
Различие транскрипционных факторов, необходимых для дифференцировки Th1- и Th2-клеток, становится особенно четким при сопоставлении структуры промоторных участков генов IFNG и IL4, кодирующих ключевые для развития этих клеток цитокины (см. рис. 3.97).
Основной источник цитокинов, обеспечивающих дифференцировку Th1-лимфоцитов, — дендритные клетки. Они выделяют IL-12 уже при первом контакте с CD4+ Т-клетками. При взаимодействии с активированными ТМ-лимфоцитами дендритные клетки секретируют весь набор цитокинов, необходимых для развития TM-клеток: IL-12, IFNy, IL-18, IL-23, IL-27 и IFNa/p. В качестве дополнительного источника IFNy выступают субпопуляции естественных киллеров и NKT-клеток. Вопрос о цитокиновом обеспечении дифференцировки ^2-клеток более сложен, потому что источники IL-4 — единственного цитокина-индуктора ^2-клеток — до их появления в ходе иммунного ответа точно не установлены. Конститутивная выработка этого цитокина в той или иной степени свойственна тучным клеткам, NK-, NKT-клеткам, эозинофилам, базофилам, но четко не показано, в какой степени она реализуется при иммунном ответе in vivo и какова доступность выделяемого IL-4 для TM-клеток. Существует предположение, что именно IL-4 служит фактором, в конечном счете определяющим баланс между Th1- и ^2-клетками, поскольку его эффект превосходит действие других цитокинов и направляет дифференцировку в сторону ^2-клеток.
К нецитокиновым факторам, влияющим на направление дифференци- ровки Т-хелперов, относят гормоны и другие медиаторы: дигидроэпианд- ростерон и релаксан индуцируют в основном TW-клетки, а глюкокортикоиды, простагландин E2, гистамин, прогестерон, дигидрооксивитамин D — ^2-лимфоциты.
Определение мембранных маркеров редко используют для дифференцирования Th1- и ^2-клеток. К таким маркерам прежде всего относят хемокиновые рецепторы: для Th1 характерны CXCR3 и CCR5, реагирующие соответственно на IFNy-зависимые a-хемокины (IP-10, MIG, I-TAC) и воспалительные р-хемокины (MIP-1a, MIP-1P, RANTES). Для ^2-кле- ток свойственны р-хемокиновые рецепторы — CCR3 (связывает эотаксин, МСР2, МСР3, МСР4 и RANTES), CCR4 (связывает TARC и MDC) и CCR8 (специфичен для I-309 и MIP-1P). Th1- и ^2-клетки различаются также по спектру и составу цитокиновых рецепторов. Так, рецептор для IL-1 1-го типа (IL-1R1) экспрессирован на Th2, но не на TM-клетках. Об особенностях экспрессии этими клетками рецепторов для IL-12 и IFNy говорилось выше. Некоторые мембранные молекулы, характерные для рассмотренных субпопуляций Т-хелперов, перечислены в табл. 3.9.
Главным подходом при разграничении Th1- и ^2-клеток служит оценка спектра секретируемых ими цитокинов, что представляется вполне адекватным, поскольку именно цитокины определяют специфические функции этих клеток. Так, ключевой цитокин TM-клеток IFNy реализует основной эффект TW-клеток — активацию макрофагов, одновременно подавляя развитие и активность ^2-клеток. IL-2, при всей важности для функционирования Thl-клеток (регуляция роста Т-клеток и активации/дифферен- цировки различных типов цитотоксических клеток), не служит для них ключевым цитокином. Более того, со временем уровень его секреции снижается (отчасти под влиянием экспрессии T-bet). Лимфотоксин а является продуктом TW-клеток, но его роль в реализации функции TW-лимфоцитов до конца не выяснена. TNFa вносит значительный вклад в проявление активности TW-клеток, особенно в комбинации с IFNy, но его выработка не является строго специфичной для этих клеток.
Функции ^2-клеток в значительной степени определяются секрецией IL-4. Наиболее важные эффекты этого цитокина — обеспечение пролиферации активированных В-клеток, ведущая роль в переключении изотипов антител на IgG1 (у мышей) и особенно на IgE, а также участие в развитии тучных клеток. К ключевым цитокинам ^2-клеток относят также IL-13 и IL-5. IL-13 воспроизводит большую часть эффектов IL-4; кроме того, он стимулирует продукцию слизи эпителиоцитами и отвечает за перестройку эпителия слизистых оболочек. IL-5 обепечивает развитие эозинофилов и служит для них хемоаттрактантом, а также участвует в поддержании пролиферации В-клеток. IL-10 в большей степени специфичен для ^2-клеток мыши, чем человека. Этот цитокин играет важную роль в запуске образования антител и ингибировании TM-клеток. Более детальная характеристика Th1- и ^2-цитокинов представлена ниже.
Помимо свойств цитокинов, вырабатываемых Th1- и ^2-клетками, их функция зависит от прямых контактных взаимодействий с клетками-мишенями, выступающими в качестве исполнителей эффекторных функций при иммунном ответе. Для TW-клеток такими мишенями служат макрофаги, для ^2-клеток — В-лимфоциты. Сигналы, индуцируемые под влиянием контактных и гуморально опосредованных воздействий со стороны Т-хелперов, определяют вклад этих лимфоцитов в развитие соответственно клеточного (в его воспалительном варианте) и гуморального иммунного ответа.
Через цитокины Th1- и ^2-клетки способны ингибировать развитие и функционирование друг друга. Уже отмечалось, что IFNy, продуцируемый TM-лимфоцитами, подавляет развитие ^2-клеток; у мышей подавление развития TW-лимфоцитов реализуются преимущественно при помощи IL-10, а у человека в большей степени — через IL-4.
- Th1- и ТЬ2-клетки
Дихотомию Т-хелперного звена системы адаптивного иммунитета впервые описали в 1986 г. Т. Мосманн (T.R. Mosmann) и соавт. Они обнаружили, что панель Т-хелперных клонов мышей по способности секретировать цитокины можно разделить на 2 группы, и обозначили их как Th1 и Th2.
Таблица 3.25. Адаптивные субпопуляции Т-хелперов
|
Показатель |
Th1 |
Th2 |
Th17 |
|
Типичные индукторы |
Внутриклеточные патогены (например, микобактерии) |
Паразиты, аллергены |
Внеклеточные патогены |
|
Факторы, благоприятствующие развитию |
Высокие и низкие дозы антигены, релаксан, дигид- роэпиандростерон, полный адъювант Фрейнда |
Промежуточные дозы антигена, глюкокортикоиды, простагландин Е2, гистамин, прогестерон, дигидрооксивитамин D, алюминиевые квасцы |
Презентация антигена дендритными клетками и макрофагами, стимулированными через TLR или CD40 |
|
Цитокины- индукторы |
IL-12, IFNy, IL-18, IL-23, IL-27 |
IL-4 |
IL-6, IL-23, TGFp |
|
Дифферен- цировочные факторы |
Tbet |
GATA-3 |
ROR-C |
|
Продуцируемые цитокины |
IFNy, IL-2, TNFa, TNFp |
IL-4, IL-5, IL-13, IL-6, IL-9, IL-10 |
IL-17, IL-22, IL-21 |
|
Клетки — функциональные партнеры |
Макрофаги |
В-клетки, эозинофилы |
Нейтрофилы |
|
Защитная функция |
Защита от внутриклеточных патогенов, локализующихся в цитоплазме |
Защита от паразитов и внеклеточных патогенов |
Защита от внеклеточных патогенов |
|
Повреждающая роль |
Развитие клеточных аутоиммунных процессов |
Развитие аллергии |
Развитие аутоиммунных процессов |
Первые секретировали IFNy, IL-2, IL-3, GM-CSF, вторые — IL-4, IL-5 и IL-3. Позже дифференцировка Т-хелперов на 2 субкласса была показана in vivo при иммунном ответе и была продемонстрирована функциональная значимость Th1/Th2-дихотомии. Этот важный вывод был сделан при анализе природы различий в чувствительности к Leishmania major мышей линий BALВ/c (чувствительная линия) и C57BL/6 (устойчивая линия). Оказалось, что у мышей C57BL/6 в ответ на инфицирование лейшманиями формируются преимущественно Th1-, а у мышей BALB/c — ^2-клетки. Различие спектра гуморальных продуктов, прежде всего ключевых цитокинов Th1- и ^2-клеток (соответственно IFNy и IL-4) определяет основную направленность иммунного ответа в направлении клеточной или гуморальной защиты. Клеточный иммунный ответ обусловливает эффективную защиту от внутриклеточных патогенов (к которым относят лейшмании), а гуморальный ответ — защиту от внеклеточных патогенов. Только соответствие направления дифференцировки Т-хелперов (а следовательно, типа ответа) природе патогена делает эту защиту эффективной. Поэтому мыши линии C57BL/6, формирующие Th1-зависимый клеточный ответ, эффективно защищаются от лейшманиазной инфекции, а мыши линии BALB/c, реагирующие развитием ^2-зависимого гуморального ответа, оказываются беззащитными и гибнут.
Открытие адаптивных субпопуляций Т-хелперов создало основу для формирования представлений о различии клеточных механизмов иммунной защиты, в зависимости от локализации патогена (внутри- или внеклеточная). Это сильно повлияло также на представления о синтезе цитокинов (двойственность набора цитокиновых продуктов показана не только для Т-хелперов, но и для ряда других клеток иммунной системы) и послужило толчком для изучения путей дифференцировки Т-хелперов. Наконец, это открытие оказало значительное влияние на понимание природы таких форм иммунопатологии, как аллергия и аутоиммунные процессы, в развитии которых важную роль играет дисбаланс субпопуляций Т-хелперов. При всей важности этой концепции необходимо сознавать, что она в значительной степени схематизирует иммунологические процессы, что особенно заметно проявляется при рассмотрении иммунных процессов, происходящих в организме человека.
При дифференцировке Т-хелперов происходит супрессия одних и усиление экспрессии других цитокиновых генов, прежде слабо экспрессированных в ThO-клетках. ТЫ-клетки продуцируют IFNy, IL-2, TNFa, TNFp, IL-3, GM-CSF, Th2-клетки - IL-4, IL-13, IL-5, IL-6, IL-9, IL-11, IL-21, IL-25, IL-10, IL-3, GM-CSF. В этих спектрах есть цитокины, общие для двух линий (GM-CSF, IL-3, у человека — IL-10). Другие цитокины более или менее специфичны для субпопуляций. Среди этих цитокинов выделяют ключевые для Th1- и ^2-клеток, поскольку они не просто специфичны для них, но и причастны к выполнению их основных эффекторных функций, индукции дифференцировки клеток этих субпопуляций, самоподдержанию и подавлению развития клеток противоположного типа. Для ТЫ-клеток таким цитокином является IFNy, для ^2-клеток — IL-4 (рис. 3.100).
Факторы и механизмы дифференцировки Th1- и ^2-клеток изучены достаточно детально. Их дифференцировка зависит от действия по меньшей мере двух сигналов. Один из них поступает через TCR и «сообщает» о распознавании антигена в комплексе с молекулой MHC. Второй сигнал поставляют цитокины. Относительная роль этих сигналов до конца не определена. Цитокинам приписывают селективную или инструктивную функции. Согласно селективной точке зрения, направление дифференци- ровки Т-хелперов задается при передаче сигнала через TCR, а цитокины только выбирают ту или иную субпопуляцию, обеспечивая ее факторами выживания и пролиферации. Согласно инструктивной точке зрения, через TCR передается активационный сигнал, тогда как направление дифферен- цировки задают цитокины через соответствующие рецепторы. В настоящее время предпочтение отдают первой точке зрения.
Развитию ТЫ-клеток способствуют крайние (очень высокая и очень низкая) дозы антигена и его высокое сродство к рецептору, а развитию ^2-клеток — промежуточные дозы антигена и более низкое его сродство
Рис. 3.100. Дифференцировка Т-хелперов типов Th1 и Th2. Указаны пути диффе- ренцировки Т-хелперов двух основных типов, функции образующихся клеток, а также цитокины, направляющие эту дифференцировку и секретируемые самими Т-хелперами
к рецептору. О значении корецептора CD4 в выборе пути дифференциров- ки свидетельствует усиление дифференцировки Т-хелперов в направлении TM-клеток при нокауте гена CD4. С решающей ролью в дифференциров- ке субпопуляций Т-хелперов антигенного сигнала можно связать то, что способность вызывать разные формы иммунного ответа через индукцию ТЫ- или ^2-клеток обусловлена самой природой антигенов. Так, преимущественную дифференцировку TM-хелперов вызывают антигены лейшма- ний, трипаносом, хламидий, микобактерий, бордетелл, бореллий, гелико- бактера, кандид, вируса гриппа, а также бактериальная ДНК, содержащая последовательность CpG, двуспиральная РНК, ЛПС и суперантигены. Избирательными индукторами ^2-хелперов служат антигены шистосом, токсокар, вирусов оспы, кори, а также аллергены и протеазы паразитов. Перечень антигенов-индукторов ТЫ-клеток наводит на мысль, что за TM-ориентацию иммунного ответа отвечают не антигены как таковые, а PAMP, с которыми они связаны. Антигенные эпитопы и детерминанты PAMP могут располагаться на одной молекуле, как например в случае ЛПС. Сигналы от PAMP могут воспринимать рецепторы АПК, прежде всего дендритных, в значительной степени определяющие характер передачи сигнала через антигенраспознающие рецепторы лимфоцитов. В случае антигенов, индуцирующих преимущественно ^2-ответ, молекулярные основы данного предпочтения не установлены. Таким образом, роль антигенов в выборе пути дифференцировки Т-хелперов скорее всего связана с передачей сигнала не через TCR, а через костимулирующие молекулы. Вероятно, влиянием на АПК можно объяснить различное действие адъювантов на диф- ференцировку Т-хелперов. Так, полный адъювант Фрейнда (содержащий микобактерии) способствует развитию Thl-клеток, а алюминиевые квасцы и столбнячный анатоксин — дифференцировке по ^2-пути.
Роль костимуляции, осуществляемой АПК, в выборе пути дифференци- ровки Т-хелперов проявляется при анализе на уровне конкретных костиму- лирующих молекул. Так, преобладающее участие в костимуляции молекул Т-клетки ICOS и OX40, а также молекулы CD86, экспрессированной на дендритной клетке, направляет дифференцировку в сторону ^2-клеток, в то же время костимуляция через CD80 не оказывает существенного влияния на дифференцировку. Предварительная обработка дендритных клеток IFNy или IL-12 придает им способность преимущественно индуцировать ТЫ-дифференцировку, а обработка IL-10 или фактором TSLP — развитие ^2-клеток. Миелоидные дендритные клетки имеют большую способность к индукции дифференцировки Thl-лимфоцитов, а плазмоцитоидные — ^2-клеток. В связи с этим сформировались представления о двух субпопуляциях дендритных клеток — DC1 и DC2, индуцирующих развитие соответственно Th1- и ^2-клеток (см. раздел 2.1.6).
Роль цитокинов в обеспечении дифференцировки Th1- и ^2-клеток изучена более детально. К настоящему времени установлено, что основным индуктором Thl-клеток служит IL-12. Развитию и поддержанию Thl-ответа способствуют также IFNy, IL-18, IL-23 и IL-27. Аналогичную, но второстепенную роль играют TNFa, TGFp, IFNa. Цитокином, определяющим развитие ^2-клеток, служит только IL-4. Роль второстепенных факторов при их развитии играют IL-10, IL-6, IL-2, IL-19 и даже цепь р40 IL-12. Th0-клетки экспрессируют рецепторы для названных цитокинов. Они несут «полные» рецепторы для IL-12 (PiP2), IFNy (aP) и IL-4 (ay), через которые получают сигналы, определяющие дальнейшую судьбу Т-хелпера. Уже после дифференцировки на некоторых клетках структура названных рецепторов становится неполной. Так, ^2-клетки утрачивают р2-цепь рецептора для IL-12. Полный рецептор для IFNy сохраняют только ^2-клетки, тогда как Th1-лимфоциты утрачивают р-цепь.
Влияние IL-12 на дифференцировку Th1-клеток реализуется через рецептор, содержащий 2 полипептидные цепи — Pj и р2. Цитоплазматические участки цепей связаны с тирозинкиназами семейства Jak: Pj-цепь связана с киназой Tyk2, а р2-цепь — с киназой Jak2. Эти киназы фосфорилируют (и, следовательно, активируют) транскрипционные факторы STAT1, STAT3, STAT4 и STAT5. Показано, что наиболее важен для дальнейшей передачи сигнала тандем р2-цепи с киназой Jak2. Именно он отвечает за фосфорилирование транскрипционного фактора STAT4, которому принадлежит ключевая роль в передаче сигнала, приводящего к дифференцировке Thl-клеток. В активации STAT4 задействован также сигнальный путь, реализуемый через MAP-киназу р38, которая фосфорилирует STAT4. Среди эффектов STAT4 следует выделить его способность взаимодействовать с промотором гена, кодирующего IFNy, и запускать его экспрессию.
Другой транскрипционный фактор, играющий ключевую роль в формировании Thl-фенотипа — T-bet (рис. 3.101). Его экспрессия коррелирует с продукцией IFNy. Экспрессия этого фактора зависит от сигналов, поступающих от TCR и IFNyR, но не зависит от сигналов, индуцируемых IL-12. Нокаут гена T-bet приводит к полному подавлению выработки IFNy и нарушению дифференцировки Th1-клеток при активации in vitro и in vivo. Выработка ^2-цитокинов при этом усиливается. Таким образом, IFNy усиливает экспрессию T-bet и в то же время экспрессия IFNy зависит от T-bet. Формируется петля положительной обратной связи и достигается стабилизация фенотипа Th1-клеток. В дифференцировке TM-клеток участвуют еще несколько факторов — Hlx, ERM, IRF-1.
Основные транскрипционные факторы, определяющие дифференци- ровку ^2-клеток, — STAT6 и GATA-3 (последнему принадлежит ведущая роль) (см. рис. 3.101). STAT6 активируется при связывании IL-4 со своим рецептором. Активация GATA-3 зависит как от сигнализации через TCR, так и от активности STAT6. GATA-3 «включает» петлю аутоактивации, которую рассматривают как механизм стабилизации фенотипа ^2-клеток. Совместно с дополнительными факторами, в частности сMaf и NF-AT, экспрессируемыми при участии STAT6, GATA-3 индуцирует экспрессию генов IL4, IL5 и IL13. В опытах с нокаутом гена GATA-3 показана однозначная связь индукции ^2-цитокинов с экспрессией GATA-3. Кроме того, GATA-3 супрессирует развитие Th1-клеток, подавляя экспрессию генов STAT4 и IL-12RP2, что затрудняет передифференцировку ^2-лимфоцитов в Th1. GATA-3 участвует в поддержании пролиферации ^2-клеток. Таким
Рис. 3.101. Вклад цитокинов и дифференцировочных факторов в образование Th1- и 1Ь2-лимфоцитов. Отражены основные внеклеточные и внутриклеточные факторы, определяющие дифференцировку Т-хелперов и их взаимное подавление
образом, фактор GATA-3 важен не только для дифференцировки ТИ2-кле- ток, но и для стабилизации их фенотипа, а также подавления программы дифференцировки TW-лимфоцитов.
Различие транскрипционных факторов, необходимых для дифференцировки Th1- и Th2-клеток, становится особенно четким при сопоставлении структуры промоторных участков генов IFNG и IL4, кодирующих ключевые для развития этих клеток цитокины (см. рис. 3.97).
Основной источник цитокинов, обеспечивающих дифференцировку Th1-лимфоцитов, — дендритные клетки. Они выделяют IL-12 уже при первом контакте с CD4+ Т-клетками. При взаимодействии с активированными ТМ-лимфоцитами дендритные клетки секретируют весь набор цитокинов, необходимых для развития TM-клеток: IL-12, IFNy, IL-18, IL-23, IL-27 и IFNa/p. В качестве дополнительного источника IFNy выступают субпопуляции естественных киллеров и NKT-клеток. Вопрос о цитокиновом обеспечении дифференцировки ^2-клеток более сложен, потому что источники IL-4 — единственного цитокина-индуктора ^2-клеток — до их появления в ходе иммунного ответа точно не установлены. Конститутивная выработка этого цитокина в той или иной степени свойственна тучным клеткам, NK-, NKT-клеткам, эозинофилам, базофилам, но четко не показано, в какой степени она реализуется при иммунном ответе in vivo и какова доступность выделяемого IL-4 для TM-клеток. Существует предположение, что именно IL-4 служит фактором, в конечном счете определяющим баланс между Th1- и ^2-клетками, поскольку его эффект превосходит действие других цитокинов и направляет дифференцировку в сторону ^2-клеток.
К нецитокиновым факторам, влияющим на направление дифференци- ровки Т-хелперов, относят гормоны и другие медиаторы: дигидроэпианд- ростерон и релаксан индуцируют в основном TW-клетки, а глюкокортикоиды, простагландин E2, гистамин, прогестерон, дигидрооксивитамин D — ^2-лимфоциты.
Определение мембранных маркеров редко используют для дифференцирования Th1- и ^2-клеток. К таким маркерам прежде всего относят хемокиновые рецепторы: для Th1 характерны CXCR3 и CCR5, реагирующие соответственно на IFNy-зависимые a-хемокины (IP-10, MIG, I-TAC) и воспалительные р-хемокины (MIP-1a, MIP-1P, RANTES). Для ^2-кле- ток свойственны р-хемокиновые рецепторы — CCR3 (связывает эотаксин, МСР2, МСР3, МСР4 и RANTES), CCR4 (связывает TARC и MDC) и CCR8 (специфичен для I-309 и MIP-1P). Th1- и ^2-клетки различаются также по спектру и составу цитокиновых рецепторов. Так, рецептор для IL-1 1-го типа (IL-1R1) экспрессирован на Th2, но не на TM-клетках. Об особенностях экспрессии этими клетками рецепторов для IL-12 и IFNy говорилось выше. Некоторые мембранные молекулы, характерные для рассмотренных субпопуляций Т-хелперов, перечислены в табл. 3.9.
Главным подходом при разграничении Th1- и ^2-клеток служит оценка спектра секретируемых ими цитокинов, что представляется вполне адекватным, поскольку именно цитокины определяют специфические функции этих клеток. Так, ключевой цитокин TM-клеток IFNy реализует основной эффект TW-клеток — активацию макрофагов, одновременно подавляя развитие и активность ^2-клеток. IL-2, при всей важности для функционирования Thl-клеток (регуляция роста Т-клеток и активации/дифферен- цировки различных типов цитотоксических клеток), не служит для них ключевым цитокином. Более того, со временем уровень его секреции снижается (отчасти под влиянием экспрессии T-bet). Лимфотоксин а является продуктом TW-клеток, но его роль в реализации функции TW-лимфоцитов до конца не выяснена. TNFa вносит значительный вклад в проявление активности TW-клеток, особенно в комбинации с IFNy, но его выработка не является строго специфичной для этих клеток.
Функции ^2-клеток в значительной степени определяются секрецией IL-4. Наиболее важные эффекты этого цитокина — обеспечение пролиферации активированных В-клеток, ведущая роль в переключении изотипов антител на IgG1 (у мышей) и особенно на IgE, а также участие в развитии тучных клеток. К ключевым цитокинам ^2-клеток относят также IL-13 и IL-5. IL-13 воспроизводит большую часть эффектов IL-4; кроме того, он стимулирует продукцию слизи эпителиоцитами и отвечает за перестройку эпителия слизистых оболочек. IL-5 обепечивает развитие эозинофилов и служит для них хемоаттрактантом, а также участвует в поддержании пролиферации В-клеток. IL-10 в большей степени специфичен для ^2-клеток мыши, чем человека. Этот цитокин играет важную роль в запуске образования антител и ингибировании TM-клеток. Более детальная характеристика Th1- и ^2-цитокинов представлена ниже.
Помимо свойств цитокинов, вырабатываемых Th1- и ^2-клетками, их функция зависит от прямых контактных взаимодействий с клетками-мишенями, выступающими в качестве исполнителей эффекторных функций при иммунном ответе. Для TW-клеток такими мишенями служат макрофаги, для ^2-клеток — В-лимфоциты. Сигналы, индуцируемые под влиянием контактных и гуморально опосредованных воздействий со стороны Т-хелперов, определяют вклад этих лимфоцитов в развитие соответственно клеточного (в его воспалительном варианте) и гуморального иммунного ответа.
Через цитокины Th1- и ^2-клетки способны ингибировать развитие и функционирование друг друга. Уже отмечалось, что IFNy, продуцируемый TM-лимфоцитами, подавляет развитие ^2-клеток; у мышей подавление развития TW-лимфоцитов реализуются преимущественно при помощи IL-10, а у человека в большей степени — через IL-4.
Источник: Ярилин.А.А , «Иммунология » 2010
А так же в разделе « Дифференцировка Т-хелперов »
- Презентация антигена
- Миграция клеток, участвующих в презентации антигена
- Иммунный синапс
- Костимуляция
- Активация Т-лимфоцитов
- Пролиферативная экспансия клонов Т-хелперов
- Th17 и другие адаптивные субпопуляции Т-клеток
- ИММУННЫЙ ОТВЕТ
- Клеточный иммунный ответ
- Воспалительный Т-клеточный иммунный ответ
- Гуморальный иммунный ответ
- Активация В-лимфоцитов. Роль Т-клеток и цитокинов
- Дифференцировка и селекция В-клеток в зародышевых центрах
- Гибридомы и моноклональные антитела. Генно-инженерные антитела
- Иммунологическая память и вторичный иммунный ответ
- В-клетки памяти
- Т-клетки памяти
- Вторичный иммунный ответ
- Неклассические проявления иммунных реакций
- 3.6.4.3. Проявления активности у5Т- и CD8aa+ Т-клеток
- Иммунологические функции NKT-клеток
- Иммунные процессы в слизистых оболочках (мукозальный иммунный ответ)
- Афферентное и центральное звенья мукозального иммунного ответа
- Роль миграции клеток в мукозальном иммунитете
- Эффекторные механизмы мукозального иммунитета
- Контроль и регуляция иммунного ответа
- Генетический контроль иммунного ответа
- Эндокринный и нервный контроль иммунного ответа
- Регуляция иммунного ответа
- Регуляторные Т-клетки
- РЕЗЮМЕ