Неклассические проявления иммунных реакций
Главные участники и исполнители адаптивного иммунитета — Т- и В-лимфоциты с их вариабельными рецепторами и клональной структурой популяций. NK-клетки, напротив, лишены клональной варибельности и функционируют в рамках врожденного иммунитета, хотя и отличаются рядом признаков от классических представителей этой ветви иммунитета — миелоидных клеток. Существует несколько субпопуляций В- и Т-лимфоцитов, занимающих промежуточное положение между врожденным и адаптивным иммунитетом. Они несут вариабельные рецепторы для антигена, но их вариабельность выражена слабее, чем у классических Т- и В-лимфоцитов. В то же время эти клетки имеют свойство клеток врожденного иммунитета — готовность к выполнению защитных функций без предварительной, достаточно длительной фазы подготовки.
Знания об этих субпопуляциях пока очень ограничены и их место в иммунной системе определено не четко. Несмотря на клональную структуру и способность распознавать индивидуальные антигены, эти клетки тем не менее по особенностям реактивности следует относить к клеткам врожденного иммунитета. Придерживаемся компромиссного варианта и рассматриваем эти клетки в рамках адаптивного иммунитета в качестве клеток «промежуточного типа» (эквивалент англ. от термина Innate-like) (рис. 3.127).
- Функциональная активность В1-клеток
По своим функциональным характеристикам В1-клетки (см. раздел 3.3.1.3) принадлежат к группе «промежуточных» лимфоидных клеток. B1-лимфоциты отличаются от обычных В2-клеток особенностями локализации и развития. Большинство В1-лимфоцитов отвечают на ТН-анти- гены, т.е. не требуют обязательного участия в ответе Т-лимфоцитов. Есть
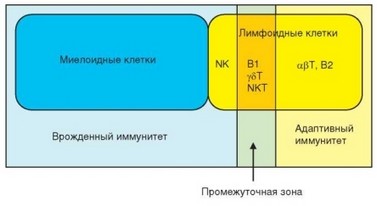
Рис. 3.127. Миелоидные и лимфоидные клетки «обслуживают» различные типы иммунитета. Цвет овалов соответствует двум основным типам иммуноцитов, фоновый цвет прямоугольников — типам иммунитета. Следует обратить внимание на промежуточную зону, в которой специфическое распознавание антигена антигенраспознающими рецепторами сочетается с быстротой мобилизацией клеток и независимостью реакции от иммунного ответа
данные, что В1-клетки способны продуцировать антитела конститутивно, т.е. независимо от стимуляции антигеном. Однако в этом вопросе много неясного, поскольку нельзя исключить активацию этих клеток аутоантигенами. К этому следует добавить, что подавляющее большинство В1-клеток в брюшной полости (основном месте их локализации) имеют активированный фенотип (CD80/86+), т.е. несут следы предшествовавшего «антигенного опыта».
Природа стимулов, запускающих дифференцировку В1-клеток в антитело-продуценты, до конца невыяснена. В1-клетки функционируют преимущественно в lamina propria слизистых оболочек и селезенке. В основных местах их локализации — в серозных полостях — В1-лимфоциты, по-видимому, не продуцируют иммуноглобулины. Причина этого в настоящее время неизвестна, но, вероятно, угнетение активности В-клеток в серозных полостях обусловлено действием локального микроокружения. В1-клетки не мигрируют в первичные фолликулы и зародышевые центры вторичных фолликулов в силу отсутствия у них рецептора CXCR5.
Поскольку В1-лимфоциты отвечают преимущественно на ТН-антигены, в процессе их развития не происходит переключения изотипов, гипермутагенеза и созреванаия аффинитета. Исключение составляют IgA-продуценты слизистой оболочки кишечника (до 50% из них происходит от В1-лимфо- цитов). Поскольку В1-клетки экспресиируют молекулы CD80 и OD86, они могут выступать в качестве АПК. Таким образом, несмотря на то, что для ответа на антигены В1-клеткам непосредственная помощь Т-лимфоцитов не требуется, сами В1-клетки могут индуцировать дифференцировку наивных CD4+ Т-клеток (в Th1- и ThH-клетки, но не регуляторные Т-лимфоциты).
В1-клетки продуцируют антитела в основном классов IgM или IgA. V-домены этих антител кодируются зародышевыми последовательностями генов. Эти антитела обладают низким сродством к антигену, для них характерна полиспецифичность. Обычно они взаимодействуют с аутоантигенами (ДНК, компоненты цитоскелета, фосфатидилхолин и т.д.) или распространенными антигенами микроорганизмов (например, полисахаридами клеточной оболочки бактерий). Последнее обстоятельство, а также спонтанный (независмый от поступления антигена) синтез иммуноглобулинов делают эти клетки важным компонентом врожденного иммунитета. Эта их функция реализуется следующим образом. В организме присутствуют пресинтезированные малоспецифичные (полиспецифичные) низкоаффинные антитела (нормальные иммуноглобулины). При проникновении патогена во внутреннюю среду организма эти антитела реагируют с антигенами его поверхности. Сами по себе такие антитела не могут повредить микроорганизм. Они также не способны привлекать через Fc-рецепторы макрофаги и другие эффектор- ные клетки, поскольку рецепторов для Fc-части IgM не существует. Однако иммунные комплексы, образуемые этими антителами, связывают комплемент и обусловливают опсонизацию патогена фрагментами C3b и C3d, что облегчает его фагоцитоз. Несмотря на слабую аффинность антител, продуцируемых В1-клетками (и, следовательно, неустойчивость их связывания с патогенами), их защитная роль достаточно велика, поскольку она реализуется в самый ранний срок после инфицирования, когда более совершенные опсони- зирующие средства отсутствуют (пентраксины, выполняющие аналогичную роль, обладают еще меньшей специфичностью и сродством к патогенам).
- Тимуснезависимый иммунный ответ и антигеннезависимая дифферен- цировка антителообразующих клеток
Гуморальный ответ на ТН-антигены существенно отличается от классического тимусзависимого ответа. При этом сильно различается ответ на T^1- и TН-2-антигены (о тимуснезвисимых антигенах см. раздел 3.2.1.2).
Как известно, ТН2-антигены — полимеры (обычно полисахаридные) с повторяющимися идентичными эпитопами (например, конъюгаты динитрофенола с декстраном или фиколлом). Это позволяет одной молекуле антигена связываться с многими BCR и вызвать их кластеризацию, что служит достаточным стимулом для активации В-клеток без дополнительных сигналов, поставляемых Т-клетками через костимулирующие молекулы CD40. С другой стороны, полисахаридные антигены не презентируются Т-лимфоцитам, что исключает возможность запуска ими Т-зависимого иммунного ответа. В то же время показано, что В-клетки при ответе на ТН-2 -антигены получают сигнал через CD40. Однако в этом участвует не CD154 (CD40L), а какая-то другая молекула, вероятно, экспрессируемая не Т-клеткой. На такое воздействие способны реагировать только зрелые В-клетки. Источниками цитокиновых сигналов, поддерживающих пролиферацию этих клеток, служат NK-клетки, макрофаги или Т-лимфоциты (при классическом ответе, опосредованном В2-клетками, источником таких сигналов служат преимущественно ^2-лимфоциты).
Ответ на ТН-2-антигены проходит вне зародышевых центров, что определяет его недостатки: слабовыраженное переключение изотипов, практически полное отсутствие гипермутагенеза и повышения аффинности, слабое развитие памяти (клетки памяти практически не образуются). Тем не менее при ответе на ТН-2-антигены образуются антитела не только IgM-, но и IgG3- и IgA-классов. В результате ответ на ТН-2-антигены существенно уступает в «качестве» тимусзависимому. В то же время в процессе дифференцировки В-лимфоцитов в плазматические клетки при ответе на ТН-2-антигены, как и при ответе на тимусзависимые антигены, экспреси- руются транскрипционный фактор Blimp-1, маркирующий этот этап дифференцировки. Поскольку распространенные компоненты многих патогенов, в особенности каспулярные антигены бактерий, обладают свойствами ТН-2-антигенов, для гуморального иммунного ответа на них характерна определенная степень дефектности, а развитие Т-клеточного иммунного ответа невозможно. Таким образом, особенности гуморального иммунного ответа на ТН-2-антигены определяются их свойствами — неспособностью стимулировать Т-клетки, частично компенсируемой способностью автономно активировать В-лимфоциты при минимальном участии вспомогательных клеток.
К перечисленным недостаткам гуморального иммунного ответа на ТН-2-антигены следует отнести также его позднее становление в онтогенезе. Это связано с упоминавшимся ранее фактом (см. раздел 3.2.1.2) — ответ на ТН-2-антигены могут осуществлять только наиболее зрелые (у мышей — Lyb5+) В-клетки, популяция которых формируется в онтогенезе только к 2—3 годам. Это обстоятельство имееет важное практическое значение, поскольку иммунизация бактериальными полисахаридами малоэффективна до указанного возраста. Эту проблему можно решить, используя полисахариды, конъюгированные с белками, что придает им свойства тимусзависи- мых антигенов.
Механизмы гуморального иммунного ответа на ТН-1-антигены изучены еще слабее. Способность ТН-1-антигенов активировать В-лимфоциты была рассмотрена в разделе З.6.4.1. Напомним, что в малых концентрациях ТН-1-антигены активируют только клетки специфических клонов. Однако при большой концентрации эти антигены приобретают митогенные свойства и становятся способны активировать все В-клетки, независимо от специфичности их BCR. Активированная В1-клетка не мигрирует в зародышевый центр и не проходит этапов «усовершенствования» (переключения изотипов, созревание аффинитета и т.д.). В этой ситуации практически отсутствует формирование клеток памяти.
В настоящее время открывают все новые факты, расширяющие наше понимание процессов, задействованных в ТН-ответе и не вполне укладывающиеся в рамки существующих гипотез. Таким образом, нельзя исключить, что многие представления о ТН-ответе претерпят значительные изменения в ближайшее десятилетие.
Давно отмечено, что В-лимфоциты, являясь типичными лимфоидными клетками и участвуя в процессах, обеспечивающих адаптивную иммунную защиту, обладают свойствами, характерными в большей степени для мие- лоидных клеток. Наиболее известный пример — принадлежность В-лимфо- цитов к АПК. В-лимфоциты (как и другие лимфоидные клетки) экспрессируют TLR (у человека — TLR-7, TLR-9, TLR-10; у мышей также TLR-4).
Это обусловливает способность В-клеток не только пролиферировать, но и дифференцироваться (поликлонально) в ответ на действие CpG-содержа- щей ДНК, а В-клеток мышей — также в ответ на ЛПС. Антигеннезависи- мую дифференцировку В-клеток вызывает действие ряда цитокинов — IL-10, IL-21 при обязательном сочетании с передачей через молекулу CD40 сигналов от Т-лимфоцитов. Вероятно, антигеннезависимая стимуляция В-клеток вносит вклад, возможно решающий, в индукцию образования при иммунном ответе антигеннеспецифических антител, доля которых может превышать долю специфических антител.
Источник: Ярилин.А.А , «Иммунология » 2010
А так же в разделе «Неклассические проявления иммунных реакций »
- Презентация антигена
- Миграция клеток, участвующих в презентации антигена
- Иммунный синапс
- Костимуляция
- Активация Т-лимфоцитов
- Пролиферативная экспансия клонов Т-хелперов
- Дифференцировка Т-хелперов
- Th17 и другие адаптивные субпопуляции Т-клеток
- ИММУННЫЙ ОТВЕТ
- Клеточный иммунный ответ
- Воспалительный Т-клеточный иммунный ответ
- Гуморальный иммунный ответ
- Активация В-лимфоцитов. Роль Т-клеток и цитокинов
- Дифференцировка и селекция В-клеток в зародышевых центрах
- Гибридомы и моноклональные антитела. Генно-инженерные антитела
- Иммунологическая память и вторичный иммунный ответ
- В-клетки памяти
- Т-клетки памяти
- Вторичный иммунный ответ
- 3.6.4.3. Проявления активности у5Т- и CD8aa+ Т-клеток
- Иммунологические функции NKT-клеток
- Иммунные процессы в слизистых оболочках (мукозальный иммунный ответ)
- Афферентное и центральное звенья мукозального иммунного ответа
- Роль миграции клеток в мукозальном иммунитете
- Эффекторные механизмы мукозального иммунитета
- Контроль и регуляция иммунного ответа
- Генетический контроль иммунного ответа
- Эндокринный и нервный контроль иммунного ответа
- Регуляция иммунного ответа
- Регуляторные Т-клетки
- РЕЗЮМЕ