
Оспо-, асфаро- и иридовирусы имеют вирионную транскриптазу (ДНК-зависимую РНК-полимеразу) и размножаются в цитоплазме. Их очень большие геномы кодируют многочисленные ферменты, которые делают их репликацию независимой от клеточного ядра. Моноцистронные мРНК транскрибируются непосредственно на вирусной ДНК.
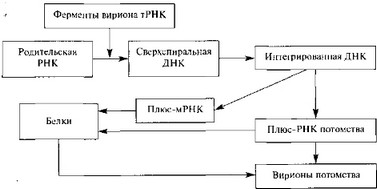
После разрушения наружной оболочки вириона клеточными ферментами на вирусной ДНК с помощью вирионной ДНК-зависимой РНК-полимеразы синтезируются ранние мРНК. Они транслируются в вирусные белки с помощью которых вирионная ДНК полностью освобождается от белка и реплицируется по полуконсервативному механизму. Вначале синтезируются фрагменты ДНК (фрагменты Оказаки), которые затем соединяются с образованием зрелых вирусных ДНК. Поздние моно- и полицистронные мРНК транскрибируются после начала репликации вирусной ДНК и кодируют синтез структурных белков вириона. Иридовирусы и асфаровирусы, по-видимому, обладают сходной с вирусами оспы стратегией репликации [286, 1109, 1135].
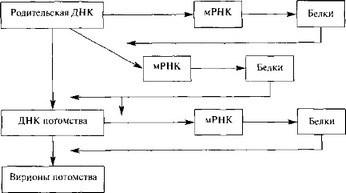
Герпес-, адено-, папиллома и полиомавирусы в одном отношении имеют общую стратегию репликации: вирусная ДНК транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. В течение определенного промежутка времени проходит два или более циклов транскрипции различных генных единиц (групп генов под контролем одного промотора). Субгеномные полицистронные РНК-транскрипты подвергаются расщеплению и сплайсингу, превращаясь в моноцистронные мРНК. У папиллома- и полиомавирусов транскрипционная программа состоит по крайней мере из двух циклов (ранние и поздние мРНК), а у герпес- (рис. 11) и аденовирусов по крайней мере из трех (сверхранние, ранние и поздние мРНК). В каждом случае вирионные полипептиды синтезируются на мРНК, появляющейся в последнем цикле транскрипции (поздние мРНК). ДНК аденовирусов, папиллома- и полиомавирусов может встраиваться в геном клетки-хозяина, реплицироваться вместе с ним, передаваться дочерним клеткам и транслироваться с образованием вирусных белков [286, 1323,1403,1135].
Парво- и цирковирусы, обладающие оцЦНК, реплицируются в ядре. Эти вирусы используют клеточные ДНК-полимеразы для синтеза двуспиральной ДНК, которая затем транскрибируется в ядре клеточной ДНК-зависимой РНК-поли- меразой II. Синтезированные транскрипты подвергаются сплайсингу и превращаются в мРНК.
Вирионная ДНК служит матрицей для синтеза (+) и (-)ДНК. Последние включаются в вирионы в качестве генома.
Различают недефектные (автономные) и дефектные парвовирусы. Первые размножаются самостоятельно, тогда как вторые реплицируются только в присутствии вируса-помощника, которым является любой аденовирус или вирус простого герпеса. В связи с этим дефектные парвовирусы называют аденоассо- циированными вирусами (ААВ). В вирионах недефектных парвовирусов содержится односпиральная линейная (+)ДНК. Дефектные парвовирусы являются уникальными в мире вирусов, поскольку комплементарные плюс- и минус-ДНК входят в различные вирионы [286, 1403, 1135].
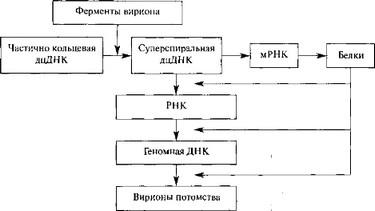
Гепаднавирусы. Частично двуцепочечная ДНК генома гепаднавирусов с помощью вирионной ДНК-полимеразы превращается в суперспирализованную двуцепочечную ДНК. Затем с помощью клеточной РНК-полимеразы II образуются мРНК для вирусных белков, в том числе для обратной транскриптазы, и полноразмерная (+)РНК, которая служит матрицей для обратной вирусной транскрипции с образованием (—)ДНК, на которой в свою очередь синтезируется частично двуцепочечная ДНК. мРНК транскрибируется на двуцепочечной ДНК, стартуя от различных промоторов (рис. 12) [693].
У оц(+)РНК-вирусов репликация начинается с полной или частичной трансляции генома без промежуточных этапов. У всех других классов вирусов процесс экспрессии вирусного генома начинается с транскрипции мРНК.
В случае с ДНК-вирусами, реплицирующимися в ядре, эту функцию выполняет клеточная ДНК-зависимая РНК-полимераза II. Все другие вирусы нуждаются в уникальной и специфичной транскриптазе, которую кодирует вирус, и она является структурным компонентом вириона. Двуцепочечные ДНК-вирусы, которые размножаются в цитоплазме, имеют ДНК-зависимую РНК-полимеразу, тогда как двуцепочечные РНК-вирусы имеют РНК-зависимую РНК-полимера- зу, специфичную для двуцепочечных РНК, а негативно полярные односпиральные РНК-вирусы носят РНК-зависимую РНК-полимеразу, специфичную для одноцепочечных РНК.
Транскрипция вирусных ДНК и РНК контролируется сложными регуляторными механизмами и, прежде всего, продуктами экспрессии регуляторных генов.
Примером некоторых механизмов регуляции экспрессии вирусных геномов, имеющих принципиальное, но не исключительное значение, могут служить аденовирусы.
У аденовирусов имеются несколько транскрипционных единиц. На различных стадиях репликативного цикла имеются «предранние», «ранние», «промежуточные» и «поздние» транскрипционные единицы, которые транскрибируются в разные периоды.
Транскрипты, которые образовались особенно в ранней стадии инфекции, считываются полностью на поздней стадии инфекции с образованием ряда длинных транскриптов с различными функциями.
Общими закономерностями транскрипции некоторых вирусов с дцДНК-ге- номом являются: транскрипция ранних и поздних генов с разных цепей ДНК, наличие перекрывающихся генов, разных рамок считывания и интронов.
У РНК-вирусов регуляция транскрипции в общем происходит менее сложно, чем у ДНК-вирусов. Временные различия транскрипции разных генов выражены не так отчетливо. У большинства семейств вирусов с оц(+)РНК геномная РНК служит мРНК, и для транскрипции-репликации РНК требуется только образование негативной цепи. Однако для ретровирусов и вирусов с несегментиро- ванным негативным РНК-геномом существут другие механизмы регуляции транскрипции.
Различные виды мРНК имеют различный период полураспада, что может также служить одним из возможных уровней регуляции вирусной репликации.
Первичные РНК-транскрипты, образующиеся на ДНК-геномах в ядре перед выходом в цитоплазму, претерпевают серию посттрансляционных изменений (кэпирование, аденилирование, метилирование, делетирование и сплайсинг). [282, 286, 1135].
Измененные вирусные моноцистронные мРНК связываются с рибосомами и транслируются в белки в основном с 5'-конца так же, как клеточные мРНК. У некоторых вирусов транскрибируется полицистронная мРНК, которая может транслироваться прямо в несколько генных продуктов. Имеется несколько механизмов перекрывания рамок считывания, что значительно расширяет кодирующий потенциал вирусов.
Это особенно важно для вирусов с небольшим геномом и еще раз указывает на экономичность экспрессии вирусных геномов, кодирующих сложные функции минимальными нуклеотидными последовательностями. Примером сказанному могут служить лентивирусы, содержащие всего 9 генов, но обладающие сложным контролем транскрипции и циклом репликации, а также удовлетворяющими требованиями установления латентной инфекции.
Большинство вирусных белков претерпевают посттрансляционные изменения, такие как: фосфолирование (для связывания нуклеиновой кислоты), ацил- лирование жирных кислот (для включения в мембрану), гликозилирование, ми- ристилированис или протсолитическос расщепление.
Вирусы используют клеточный метаболизм для синтеза боковых цепей гликопротеинов вирусной оболочки.
Посттрансляционное расщепление вирусных белков имеет место у многих вирусов, у которых полицистронная вирусная (+)РНК транслируется непосредственно в один полипротеин, который несет протеазную активность, расщепляющую полипротеин в определенных местах на мелкие белки. Первое расщепление происходит, когда полипротеин еще связан с рибосомой. Некоторые большие фрагменты существуют только быстротечно, тогда как другие функционируют короткий период и в последующем расщепляются дополнительно протеазами, кодируемыми вирусом, на мелкие белки с различными функциями.
Некоторые вирусы кодируют несколько различных протеаз, большинство из которых является трипсиноподобными. Клеточные протеазы присутствуют в таких органеллах как аппарат Гольджи или транспортные вакуоли, которые принимают участие в сборке и созревании многих вирусов. Например, расщепление гемагглютинина ортомиксовирусов или гликопротеина сплавления (F белок) па- рамиксовирусов имеет важное значение для инфекционности вирионов.
В таблице 4 приведены белки, кодируемые вирусными геномами. В общем, белки, транслируемые с ранних транскриптов ДНК-вирусов, включают ферменты и другие белки, необходимые для репликации вирусных нуклеиновых кислот белков, подавляющих синтез клеточных РНК и белков. Крупные ДНК- вирусы (оспо- и герпесвирусы) также кодируют многочисленные ферменты, связанные с метаболизмом нуклеотидов. Поздние вирусные белки транслируются на поздних мРНК, большинство из которых транскрибируется на молекулах потомства вирусных нуклеиновых кислот. Большинство поздних белков
являются вирусными структурными белками и часто образуются в избыточном количестве.
Некоторые вирусные белки обладают другими важными функциями. К ним относятся регуляторные белки, изменяющие транскрипцию или трансляцию клеточных генов или ранних вирусных генов. Крупные ДНК-вирусы также кодируют многие дополнительные белки, называемые вирокинами (подобие клеточных цитокинов), которые не регулируют цикл вирусной репликации, как таковой, но влияют на ответ хозяина на инфекцию [110, 282, 693, 1135].
После разрушения наружной оболочки вириона клеточными ферментами на вирусной ДНК с помощью вирионной ДНК-зависимой РНК-полимеразы синтезируются ранние мРНК. Они транслируются в вирусные белки с помощью которых вирионная ДНК полностью освобождается от белка и реплицируется по полуконсервативному механизму. Вначале синтезируются фрагменты ДНК (фрагменты Оказаки), которые затем соединяются с образованием зрелых вирусных ДНК. Поздние моно- и полицистронные мРНК транскрибируются после начала репликации вирусной ДНК и кодируют синтез структурных белков вириона. Иридовирусы и асфаровирусы, по-видимому, обладают сходной с вирусами оспы стратегией репликации [286, 1109, 1135].
Герпес-, адено-, папиллома и полиомавирусы в одном отношении имеют общую стратегию репликации: вирусная ДНК транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. В течение определенного промежутка времени проходит два или более циклов транскрипции различных генных единиц (групп генов под контролем одного промотора). Субгеномные полицистронные РНК-транскрипты подвергаются расщеплению и сплайсингу, превращаясь в моноцистронные мРНК. У папиллома- и полиомавирусов транскрипционная программа состоит по крайней мере из двух циклов (ранние и поздние мРНК), а у герпес- (рис. 11) и аденовирусов по крайней мере из трех (сверхранние, ранние и поздние мРНК). В каждом случае вирионные полипептиды синтезируются на мРНК, появляющейся в последнем цикле транскрипции (поздние мРНК). ДНК аденовирусов, папиллома- и полиомавирусов может встраиваться в геном клетки-хозяина, реплицироваться вместе с ним, передаваться дочерним клеткам и транслироваться с образованием вирусных белков [286, 1323,1403,1135].
Парво- и цирковирусы, обладающие оцЦНК, реплицируются в ядре. Эти вирусы используют клеточные ДНК-полимеразы для синтеза двуспиральной ДНК, которая затем транскрибируется в ядре клеточной ДНК-зависимой РНК-поли- меразой II. Синтезированные транскрипты подвергаются сплайсингу и превращаются в мРНК.
Вирионная ДНК служит матрицей для синтеза (+) и (-)ДНК. Последние включаются в вирионы в качестве генома.
Различают недефектные (автономные) и дефектные парвовирусы. Первые размножаются самостоятельно, тогда как вторые реплицируются только в присутствии вируса-помощника, которым является любой аденовирус или вирус простого герпеса. В связи с этим дефектные парвовирусы называют аденоассо- циированными вирусами (ААВ). В вирионах недефектных парвовирусов содержится односпиральная линейная (+)ДНК. Дефектные парвовирусы являются уникальными в мире вирусов, поскольку комплементарные плюс- и минус-ДНК входят в различные вирионы [286, 1403, 1135].
Гепаднавирусы. Частично двуцепочечная ДНК генома гепаднавирусов с помощью вирионной ДНК-полимеразы превращается в суперспирализованную двуцепочечную ДНК. Затем с помощью клеточной РНК-полимеразы II образуются мРНК для вирусных белков, в том числе для обратной транскриптазы, и полноразмерная (+)РНК, которая служит матрицей для обратной вирусной транскрипции с образованием (—)ДНК, на которой в свою очередь синтезируется частично двуцепочечная ДНК. мРНК транскрибируется на двуцепочечной ДНК, стартуя от различных промоторов (рис. 12) [693].
У оц(+)РНК-вирусов репликация начинается с полной или частичной трансляции генома без промежуточных этапов. У всех других классов вирусов процесс экспрессии вирусного генома начинается с транскрипции мРНК.
В случае с ДНК-вирусами, реплицирующимися в ядре, эту функцию выполняет клеточная ДНК-зависимая РНК-полимераза II. Все другие вирусы нуждаются в уникальной и специфичной транскриптазе, которую кодирует вирус, и она является структурным компонентом вириона. Двуцепочечные ДНК-вирусы, которые размножаются в цитоплазме, имеют ДНК-зависимую РНК-полимеразу, тогда как двуцепочечные РНК-вирусы имеют РНК-зависимую РНК-полимера- зу, специфичную для двуцепочечных РНК, а негативно полярные односпиральные РНК-вирусы носят РНК-зависимую РНК-полимеразу, специфичную для одноцепочечных РНК.
Транскрипция вирусных ДНК и РНК контролируется сложными регуляторными механизмами и, прежде всего, продуктами экспрессии регуляторных генов.
Примером некоторых механизмов регуляции экспрессии вирусных геномов, имеющих принципиальное, но не исключительное значение, могут служить аденовирусы.
У аденовирусов имеются несколько транскрипционных единиц. На различных стадиях репликативного цикла имеются «предранние», «ранние», «промежуточные» и «поздние» транскрипционные единицы, которые транскрибируются в разные периоды.
Транскрипты, которые образовались особенно в ранней стадии инфекции, считываются полностью на поздней стадии инфекции с образованием ряда длинных транскриптов с различными функциями.
Общими закономерностями транскрипции некоторых вирусов с дцДНК-ге- номом являются: транскрипция ранних и поздних генов с разных цепей ДНК, наличие перекрывающихся генов, разных рамок считывания и интронов.
У РНК-вирусов регуляция транскрипции в общем происходит менее сложно, чем у ДНК-вирусов. Временные различия транскрипции разных генов выражены не так отчетливо. У большинства семейств вирусов с оц(+)РНК геномная РНК служит мРНК, и для транскрипции-репликации РНК требуется только образование негативной цепи. Однако для ретровирусов и вирусов с несегментиро- ванным негативным РНК-геномом существут другие механизмы регуляции транскрипции.
Различные виды мРНК имеют различный период полураспада, что может также служить одним из возможных уровней регуляции вирусной репликации.
Первичные РНК-транскрипты, образующиеся на ДНК-геномах в ядре перед выходом в цитоплазму, претерпевают серию посттрансляционных изменений (кэпирование, аденилирование, метилирование, делетирование и сплайсинг). [282, 286, 1135].
Измененные вирусные моноцистронные мРНК связываются с рибосомами и транслируются в белки в основном с 5'-конца так же, как клеточные мРНК. У некоторых вирусов транскрибируется полицистронная мРНК, которая может транслироваться прямо в несколько генных продуктов. Имеется несколько механизмов перекрывания рамок считывания, что значительно расширяет кодирующий потенциал вирусов.
Это особенно важно для вирусов с небольшим геномом и еще раз указывает на экономичность экспрессии вирусных геномов, кодирующих сложные функции минимальными нуклеотидными последовательностями. Примером сказанному могут служить лентивирусы, содержащие всего 9 генов, но обладающие сложным контролем транскрипции и циклом репликации, а также удовлетворяющими требованиями установления латентной инфекции.
Большинство вирусных белков претерпевают посттрансляционные изменения, такие как: фосфолирование (для связывания нуклеиновой кислоты), ацил- лирование жирных кислот (для включения в мембрану), гликозилирование, ми- ристилированис или протсолитическос расщепление.
Вирусы используют клеточный метаболизм для синтеза боковых цепей гликопротеинов вирусной оболочки.
Посттрансляционное расщепление вирусных белков имеет место у многих вирусов, у которых полицистронная вирусная (+)РНК транслируется непосредственно в один полипротеин, который несет протеазную активность, расщепляющую полипротеин в определенных местах на мелкие белки. Первое расщепление происходит, когда полипротеин еще связан с рибосомой. Некоторые большие фрагменты существуют только быстротечно, тогда как другие функционируют короткий период и в последующем расщепляются дополнительно протеазами, кодируемыми вирусом, на мелкие белки с различными функциями.
Некоторые вирусы кодируют несколько различных протеаз, большинство из которых является трипсиноподобными. Клеточные протеазы присутствуют в таких органеллах как аппарат Гольджи или транспортные вакуоли, которые принимают участие в сборке и созревании многих вирусов. Например, расщепление гемагглютинина ортомиксовирусов или гликопротеина сплавления (F белок) па- рамиксовирусов имеет важное значение для инфекционности вирионов.
В таблице 4 приведены белки, кодируемые вирусными геномами. В общем, белки, транслируемые с ранних транскриптов ДНК-вирусов, включают ферменты и другие белки, необходимые для репликации вирусных нуклеиновых кислот белков, подавляющих синтез клеточных РНК и белков. Крупные ДНК- вирусы (оспо- и герпесвирусы) также кодируют многочисленные ферменты, связанные с метаболизмом нуклеотидов. Поздние вирусные белки транслируются на поздних мРНК, большинство из которых транскрибируется на молекулах потомства вирусных нуклеиновых кислот. Большинство поздних белков
|
1. |
Структурные белки вирионов (капсидные, коровые, оболочечные) |
|
2. |
Ферменты, связанные с вирионами, особенно полимеразы (транскриптазы) и др. |
|
3. |
Неструктурные белки, главным образом ферменты, необходимые для транскрипции, репликации вирусной нуклеиновой кислоты и расщепления белков |
|
4. |
Регуляторные белки, контролирующие временную последовательность экспрессии вирусного генома |
|
5. |
Белки, понижающие экспрессию клеточных генов (транскрипцию, иногда трансляцию) |
|
6. |
Онкогенные (продукты) белки и инактиваторы клеточных белков, подавляющих трансформацию клеток (герпес-, адено-, папиллома-, полиома- и ретровирусные инфекции) |
|
7. |
Белки, влияющие на вирулентность вируса, хозяинный спектр, тканевой тропизм и т.д. (наиболее сложные ДНК-вирусы) |
|
8. |
Вирокины, модифицирующие неинфицированные клетки, тем самым способствуя распространению инфекции в организме |
являются вирусными структурными белками и часто образуются в избыточном количестве.
Некоторые вирусные белки обладают другими важными функциями. К ним относятся регуляторные белки, изменяющие транскрипцию или трансляцию клеточных генов или ранних вирусных генов. Крупные ДНК-вирусы также кодируют многие дополнительные белки, называемые вирокинами (подобие клеточных цитокинов), которые не регулируют цикл вирусной репликации, как таковой, но влияют на ответ хозяина на инфекцию [110, 282, 693, 1135].