
Пикорна-, калици-, астро-, тога- и флавивирусы реплицируются наиболее коротким путем: их (+)РНК геном функционирует непосредственно как мРНК.
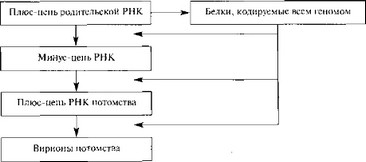
Геномы пикорна- и флавивирусов функционируют как единая полицистрон- ная мРНК, транслирующаяся прямо в единый полипротеин, который впоследствии расщепляется с образованием индивидуальных структурных и неструктурных белков. Одним из них является РНК-зависимая PH К-полимераза, которая реплицирует вирусный геном. Вирусная (+)РНК транскрибируется в комплиментарную (—)РНК-копию, которая служит матрицей для синтеза новых цепей (+)РНК (рис. 6), или может использоваться в качестве мРНК-матриц для синтеза новых минус-цепей и геномной РНК вирусного потомства.
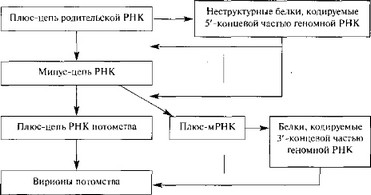
Тога-, корона- и калицивирусы отличаются от пикорнавирусов тем, что на начальном этапе инфекции экспрессируется лишь часть геномной РНК с образованием белков. Последние осуществляют синтез минус-цепи, являющейся матрицей для синтеза различных по размеру классов молекул плюс-РНК. Полипротеины, образующиеся на коротких молекулах мРНК, расщепляются на структурные вирионные белки. Полноразмерные плюс-РНК упаковываются в вирионы.
У тогавирусов транслируется только около 2/3 вирусной РНК (5'-конец); образующийся полипротеин расщепляется на неструктурные белки, которые необходимы для транскрипции и репликации РНК. Вирусная РНК-полимераза синтезирует полноразмерную (—)РНК, на которой затем синтезируются два вида (+)РНК: полноразмерная вирионная РНК, предназначенная для включения в вирионы, и РНК, длина которой равна 1/3, и которая является колинеарной с 3'-концом вирусной РНК и транслируется в полипротеин, который расщепляется на структурные белки (рис. 7). У калицивирусов образуются полигеномные и субгеномные мРНК [282, 286].
Корона- и артеривирусы демонстрируют необычную стратегию транскрипции: первоначально часть вирионной (+) РНК функционирует как мРНК и транслируется с образованием РНК-полимеразы, которая затем синтезирует полногеномную (—)РНК. На этой (—)РНК транскрибируется гнездо субгеном- ных мРНК с общими 3'-концами. Транслируются только 5'-концевые последовательности каждого члена этого гнезда транскриптов.
Главным отличием вирусов с позитивным геномом является их способность синтезировать ферменты, ответственные за репликацию вирусного генома. Поэтому РНК, выделенная из таких вирусов, инфекционна. Второе отличие состо-
ит в монолитности вирусного генома. Поэтому первичный продукт трансляции обеих РНК (геномной и мРНК) представляет собой единый белок, который в дальнейшем расщепляется на индивидуальные вирусные белки (в том числе структурные) [286].
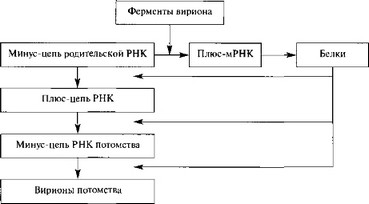
Ортомиксо-, бунья- и аренавирусы. Геном представлен (—)РНК. Каждый геномный сегмент транскрибируется отдельно вирионной РНК-транскриптазой и транслируется в один или несколько белков. Эти вирусы характеризуются тем, что их геномная РНК выполняет две матричные функции: в процессе транскрипции и репликации. Транскрипция вирусного генома — первое событие после проникновения вируса в клетку, в результате которого образуются моноцис- тронные мРНК, кодирующие один белок. Репликацию начинают вновь синтезированные вирусные белки, приводящие к образованию плюс-цепи, которая служит матрицей для синтеза геномной (—)РНК (рис. 8).
Следовательно, плюс-транскрипт, функционирующий в качестве мРНК, отличается от (+)РНК, служащей матрицей для вирусного потомства, хотя и первый, и вторая синтезируются на геномной РНК.
Следует отметить, что S-сегмент РНК аренавирусов и некоторых буньявиру- сов является двуполярным, т.е. одна часть сегмента имеет (+) полярность, другая (—) полярность. Стратегия репликации двуполярных РНК-вирусов соответствует полярности их геномов и является смешанной, присущей стратегии репликации (+)РНК и (—)РНК вирусов [286, 282, 693].
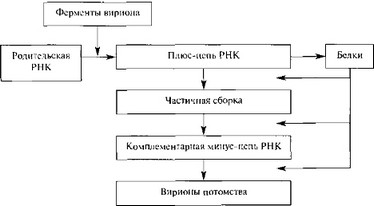
Парамиксо-, рабдо-, фило- и борнавирусы имеют геномную одноцепочечную РНК и РНК-зависимую РНК-полимеразу (транскриптазу), которая транскрибирует пять или более субгеномных (+)РНК, каждая из которых служит в каче-
стве моноцистронной мРНК. В противоположность транскрипции в результате репликации (с помощью той же самой полимеразы, действующей в качестве репликазы) образуются полноразмерные (+)цепи РНК, которые служат матрицей для синтеза новых вирионных (-)цепей РНК. Стратегия репликации борна- вирусов в некоторых деталях является более сложной [286].
Рео- и бирнавирусы имеют сегментированный двуцепочечный РНК-геном. Негативные цепи каждого сегмента (2—12 сегментов) транскрибируются раздельно в цитоплазме вирионной транскриптазой с образованием мРН К. Эти позитивно полярные (+)РНК служат матрицей для репликации. Образовавшиеся двуспиральные РНК в свою очередь служат матрицей для дальнейшей транскрипции мРНК. Транскрипция генома реовирусов происходит внутри субвирусной частицы с помощью вирионной транскриптазы, причем копируется только одна нить каждого фрагмента двуцепочечной РНК (асимметричная транскрипция). Синтезирующиеся мРНК выходят из сердцевины вириона через 12 полых вершин частично разрушенного капсида. Таким образом, молекулы мРНК реовирусов выполняют две функции. Во-первых, они транслируются, обеспечивая синтез вирусных белков, и, во-вторых, они включаются (по одной молекуле мРНК каждого из 10--12 генов) в состав частиц-предшественниц вирионов. Каждая мРНК служит в этих частицах матрицей для синтеза комплементарной цепи, что ведет к образованию двуцепочечных сегментов вирусного генома (рис. 9).
Молекулы (+)РНК, синтезированные на родительской двуспиральной РНК, связываются с белками и образуют субвирусные частицы, в которых содержится 10—12 фрагментов (+)РНК. Фрагменты (+)РНК в составе субвирусных частиц служат матрицами для синтеза (—)РНК, которые в свободном виде в инфицированных клетках не обнаружены. Полагают, что транскрипция и репликация РНК
реовирусов осуществляется одним ферментом, который модифицируется в процессе репликации вируса [164, 286, 1135].
Ретровирусы. Геном представлен двумя идентичными молекулами (+)РНК. Вместо того чтобы функционировать как мРНК, геном транскрибируется с помощью вирионной обратной транскриптазы, с образованием гибридной молекулы РНК-ДНК. Затем на (-)ДНК синтезируется (+)ДНК и образуется двуспиральная ДНК (другой активностью того же самого фермента) и непрерывно включается в клеточную ДНК. Интегрированная вирусная ДНК (провирус) транскрибируется клеточной РНК-полимеразой II с последующим сплайсингом РНК-транскриптов и расщеплением образующихся белков. Часть вновь синтезированных полноразмерных (+)РНК транскриптов соединяется в пары и образует диплоидные геномы новых вирионов (рис. 10).
Геном ретровирусов состоит из трех генов: (5' — 3') gag, pol и env, которые кодируют, соответственно, внутренние белки вируса, обратную транскриптазу и белки оболочки. Вирус саркомы Рауса имеет в своем геноме четвертый ген - онкоген (src), продукт которого играет ключевую роль в трансформации клеток, но не является необходимым для размножения вируса [110, 282, 286].