
После антигенной стимуляции, индуцирующей преимущественно клеточный иммунитет, в тимусзависимых областях лимфоидных органов наблюдается созревание и пролиферация антиген-специфических Т-лимфоцитов.
Т-эффекторы, осуществляющие клеточно-опосредованные формы иммунного ответа, характеризуются, прежде всего, цитотоксической активностью. Ци- тотоксические реакции весьма неоднородны по своим механизмам и могут опосредоваться клетками, антителами, комплементом.
Среди лимфоцитов, обладающих киллерной активностью, основную роль в элиминации вирусов из пораженных ими клеток играют вирусспецифические цитотоксические Т-лимфоциты (Тц-лимфоциты) и естественные киллеры (NK- лимфоциты). Хотя последние проявляют защитное действие на ранней стадии инфекции, тем не менее, ведущее значение в разрушении инфицированных клеток и освобождении организма от возбудителя принадлежит Тц-лимфоцитам.
Антигенспецифические Тц-лимфоциты при вирусных инфекциях взаимодействуют с клетками-мишенями и разрушают их при помощи рецепторного аппарата в отсутствии антител и комплемента. Они имеют двойную специфичность: к вирусным антигенам на поверхности инфицированных клеток и к главному комплексу гистосовместимости клеток хозяина. Для индукции Тц-клеток необходимы Т-хелперы (Тх-лимфоциты), активизированные вирусным антигеном. При ряде вирусных инфекций клетки распознают иные антигенные структуры вирусов, чем В-клетки и антитела. Это связано с рецептором Тц-клеток, распознающим вирусные антигены на клеточной поверхности, природа которого неизвестна. Следовательно, Т-клетки и антитела распознают вирусные антигены посредством различных механизмов. При этом они, как правило, распознают различные эпитопы. На молекуле НА вируса гриппа Тц-лимфоциты и антитела реагируют, соответственно, с НА2 и НА1. Вирусные антигены, являющиеся индукторами и мишенями Тц-клеток при различных вирусных инфекциях человека и животных, исследованы недостаточно. Для многих вирусов конкретные вирусные антигены, являющиеся индукторами и мишенями Тц- клеток, не идентифицированы, а имеющиеся данные зачастую противоречивы.
Основная мишень для Тц-клеток при гриппе — нуклеопротеин (NP) [181], а при РС-инфекции — белок N-вируса, тогда как гликопротеин G не распознается этими клетками. При гепатите-В основную роль в распознавании клеток-мишеней цитотоксическими клетками играет нуклеопротеин вируса, который может одновременно функционировать в качестве Т-независимого антигена. При цитомегаловирусной инфекции Т-зависимым антигеном является неструктурный вирусный белок (РР89), экспрессируемый в ранней фазе инфекции на поверхности клеток [1566]. Вирусспецифические Тц-лимфоциты лизируют инфи- цированнные ротавирусом клетки до образования инфекционного вируса, и даже выраженный синтез вирусных белков не был необходимым условием для цитолиза. Гликопротеин G вируса простого герпеса, по-видимому, является основным антигеном, ответственным за индукцию вирусспецифических Тц-лим- фоцитов. У вируса SV-40 за иммунный цитолиз ответственен Т-антиген, а у везикулярного стоматита и бешенства — белки G и N [169]. Тц-клетки обладают большей перекрестной иммунореактивностью в отношении антигенродствен- ных вирусов, чем иммуноглобулины. Например, при инфицировании мышей вирусом гриппа образуются две популяции Тц-клеток. Основная из них — типоспецифическая, способная лизировать клетки, индуцированные любым штаммом вируса гриппа А. Главным индуктором таких клеток является белок NP вируса гриппа. Штамм-специфические Тц-клетки узнавали главным образом НА- белок вируса гриппа [384]. Иммунизация мышей белком NP сопровождалась образованием перекрестно реагировавших антител и иммунитета ко всем вирусам гриппа типа А.
Перекрестная реактивность Тц-клеток обнаружена также в опытах с различными серотипами вируса катаральной лихорадки овец, ротавирусов, вируса везикулярного стоматита и альфавирусов [32].
Типоспецифический иммунитет в отношении антигеннородственных вирусов существует и в отсутствие соответствующих вируснейтрализующих антител, что свидетельствует о важной роли Тц-клеток в защите от вирусов. Широкая реактивность Тц-клеток имеет важное значение в иммунопрофилактике вирусных болезней. Продукция Тц-клеток во многом зависит от другого вирусиндуциро- ванного типа Т-клеток: Т-хелперов, которые, по-видимому, обладают сходной с Тц-клетками перекрестной реактивностью и способствуют продукции антител. Иммунитет, опосредованный Тц-клетками, играет различную роль при разных вирусных инфекциях. Считается, что индукция вирусспецифических Тц-клеток при гриппе [673] и бешенстве [50] — важный защитный компонент иммунного ответа. Адоптивный перенос Тц-лимфоцитов защищал мышей от летальной гриппозной инфекции. Тц-клетки играют важную роль в элиминации клеток, трансформированных вирусом Эпштейн-Барра [32] и вирусом болезни Марека, а также инфицированных цитомегаловирусом [15661, вирусами ветряной оспы [411] и гепатита мышей. Роль Тц-клеток в защите организма при многих вирусных инфекциях предстоит еще выяснить.
Цитотоксические клетки, зависимые от антител, вносят существенный вклад в противовирусную защиту. Они не сенсибилизированы и поэтому не могут быть отнесены к эффекторам специфического иммунного ответа.
Клеточная цитотоксичность, зависящая от антител, является своеобразным связывающим звеном между клеточным и гуморальным иммунитетом и опосредуется NK-клетками, а также нейтрофилами, несущими рецепторы к Fc-фраг- ментам иммуноглобулинов. Специфичность данного вида цитотоксичности определяется антителами, покрывающими клетки-мишени, и не зависит от комплемента.
К NK-клеткам относятся лимфоциты, не несущие ни Т-, ни В-маркеров. Поэтому их также называют «нулевыми» клетками. Специфичность лизиса инфицированных вирусом клеток определяется антителами. Этот лизис требует присутствия гораздо меньших количеств антител, чем лизис инфицированных клеток, зависимый от комплемента [107, 200]. Значение цитотоксических клеток, зависящих от специфических антител, в противовирусном иммунитете можно иллюстрировать на ряде примеров. В иммунитете при катаральной лихорадке овец важная роль принадлежит зависящим от антител цитотоксическим лимфоцитам. Гуморальные, вируснейтрализующие антитела носят типоспецифический характер, хотя между их титром и иммунитетом не всегда установлена корреляция. Важным защитным механизмом против вируса простого герпеса и болезни Ауески также является антителозависимая клеточная цитотоксичность. Однако в последнем случае выраженность вакцинального иммунитета не коррелировала с уровнем локальных или системных противовирусных антител. В некотором смысле с этими наблюдениями согласуется протективный эффект антител, не обладающих вируснейтрализующей активностью, при вирусном гепатите мышей и альфавирусной инфекции (вирус Синдбис) [1131].
NK-клетки не являются сенсибилизированными лимфоцитами и относятся к факторам неспецифической защиты, способными действовать против зараженных вирусом клеток еще до формирования специфических механизмов защиты.
NK-клетки, очевидно, представляют собой группу клеток, различающихся по способности разрушать различные клетки-мишени в зависимости от экспрессии поверхностных вирусных белков.
Пока не образовались вирусспепифические Тц- и Тх-лимфоциты и антитела, решающее значение в предотвращении и Ограничении распространения вируса в организме принадлежит клеткам, обладающим естественной киллерной активностью. Их активность в инфицированном организме быстро возрастает. Они первыми (в течение первых трех дней) вступают в борьбу, затем (через 4—7 дней) действуют Тц-клетки и антитела. Иными словами, на ранних стадиях инфекции функционируют универсальные — неспецифические защитные механизмы, позднее вступают в действие факторы специфического иммунного ответа. Иммунологические механизмы цитолиза основаны на включении вирусных белков в плазматическую мембрану клетки, которые являются мишенями для специфи-
Таблица 15. Роль клеток белой крови во врожденном и приобретенном иммунитете [670]
|
Клетки |
Источник/функция |
|
Лимфоциты |
Ответственны за специфический иммунный ответ; различают и связывают чужеродные антигены; происходят из костного мозга |
|
Т-клетки |
Различают: Т-хелперные клетки (Тх), которые секретируют цитокины; цитотоксические Т-клетки (Тц). Регуляторные или суппрессорные Т-клетки являются Тх-клетками. Все они имеют Т-клеточные рецепторы |
|
В-клетки |
Производят антитела |
|
NK-клетки |
Естественные (природные) киллерные клетки (NK): крупные гранулярные цитотоксические клетки; большинство из них не имеет Т-клеточных рецепторов |
|
Одноядерные фагоциты |
Ответственны за фагоцитоз и презентацию антигена; происходят из костного мозга; семейство моноцитов; макрофаги. Клетки Купфера в печени, альвеолярные макрофаги в легких |
|
Дендритные клетки |
Ответственны за индукцию иммунного ответа; представляют антиген Тх-клеткам; клетки мигрируют в разные ткани (органы), но не в мозг |
|
Межпальцевые дендритные клетки |
Происходят из костного мозга; присутствуют в большинстве органов, лимфоузлах и селезенке, в коже (клетки Лангерганса) |
|
Фолликулярные дендритные клетки |
Не происходят из костного мозга; находятся в герминальных слоях лимфатических фолликулов селезенки, лимфоузлов, лимфоидной ткани слизистых; являются трапом для представления В-клеткам антигенов или комплексов антиген-антитело |
|
Плазмоцитоидные дендритные клетки |
Зрелые дендритные клетки найдены в крови и Т-клеточных зонах лимфоузлов; способны синтезировать большое количество интерферона, защищающего иммунные клетки от вирусной инфекции |
|
Гранулоциты |
Содержат множество цитоплазматических гранул; клетки воспаления |
|
Нейтрофилы |
Полиморфноядерные лейкоциты; отвечающие нахемотакси- ческие сигналы; фагоцитируют чужеродные частицы; главные лейкоциты воспаления |
|
Эозлиофилы |
Функционируют в борьбе против некоторых патогенов, индуцирующих IgE; не играют важной роли в антивирусной борьбе |
|
Базофилы |
Медиаторы гиперчувствительности, вызванной IgE ответом |
ческого гуморального и клеточного иммунного ответа, следствием чего является лизис инфицированных клеток [898, 1135].
Вирусная инфекция или иммунизация могут вызвать развитие реакций гиперчувствительности замедленного типа на вирусные антигены. Это состояние сенсибилизации, выявляемое кожной пробой, опосредуется Т-клетками и представляет одну из форм клеточной иммунореактивности. Ее роль в противовирусном иммунитете еще не выяснена.
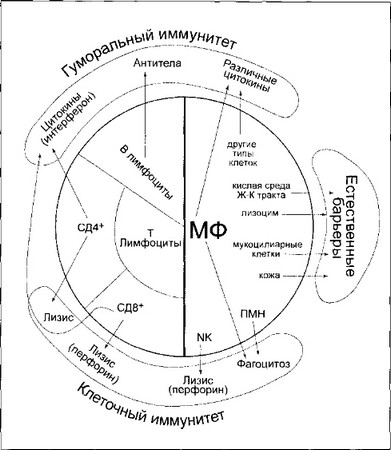
Рис. 17. Схема иммунного ответа на вирусы.
Центральная затемненная часть представляет иммунный ответ, который может быть разделен на антигенспецифические ответы (левая половина) и врожденный или неспецифический иммунитет (правая половина) [1658].
МФ-макрофаги;
NK — естественные киллерные клетки;
ПМН — полиморфноядерные нейтрофилы.
Поскольку каждый вирус обладает уникальными антигенными свойствами, роль каждого типа клеток чрезвычайно сложной иммунной системы в защите и иммунопатологии при различных заболеваниях требует индивидуальной оценки. Т-клетки, по-видимому, эффективнее всего угнетают репликацию вирусов, антигены которых экспрессируются на поверхности клеток на ранних стадиях инфекции, например, вирусов гриппа, герпес- или альфавирусов. Тц-клетки играют менее важную роль в иммунитете против тех инфекций, возбудители которых экспрессируют антигены на поверхности клеток в поздней стадии репликации (флавивирусные инфекции) или вовсе не экспрессируют их на поверхности инфицированных клеток (пикорна-, парвовирусные и другие инфекции).