
Известно, что степень защиты от локальных вирусных инфекций респираторного и желудочно-кишечного трактов прежде всего зависит от содержания в организме специфического секреторного IgA, а не от наличия сывороточного IgG к энтеропатогенным или пневмотропным вирусам [43, 139].
Стабильная структура, выраженный аффинитет к поверхности слизистых оболочек, преобладающее содержание в секрете молочной железы обусловливают биологическую роль секреторного IgA в защите организма от неблагоприятного воздействия различных патогенных агентов, в том числе вирусов.
IgA синтезируется в димерной форме в клетках lamina propria и после связывания с иммуноглобулиновым рецептором, синтезированным в эпителиальных клетках, транспортируется на поверхность слизистой оболочки. В момент выхода IgA в просвет кишечника рецептор частично расщепляется, в результате чего в составе IgA остается фрагмент рецептора, который называют секреторным компонентом (рис. 18). Таким образом, секреторный IgA является продуктом кооперации двух типов клеток — плазматических и эпителиальных.
Секреторный IgA образуется не только в димерной, но и в тетрамерной форме, что усиливает его вируснейтрализующую способность. Секреторный компонент предохраняет IgA от расщепления протеолитическими ферментами, что обусловливает его значительные преимущества перед антителами других классов. Секреторный IgA нейтрализует вирус не только в просвете кишечника, но и при транспортировке его внутрь клетки. Димер IgA может нейтрализовать вирус в подслизистой оболочке кишечника, а затем, связавшись с рецептором, транспортировать его в просвет кишечника [139].
Димерные иммуноглобулины класса А (11S IgA), соединенные посредством j-цепи в единую структуру с секреторным компонентом, представляют собой
уникальный пример эволюционной адаптации иммуноглобулинов на слизистых покровах для эффективного функционирования в условиях постоянного воздействия антигенов различной природы.
Исходя из способности к локальному синтезу молекул секреторного компонента, а также димерной формы IgA-местно расположенными плазматическими клетками, ткани слизистых покровов можно разделить на три основных класса:
Класс 1. Выраженный местный синтез IgA и секреторного компонента
Желудочно-кишечный тракт Верхний отдел респираторного тракта Носовая полость Среднее ухо Желчный пузырь
Класс 2. Местный синтез секреторного компонента при отсутствии местного синтеза IgA
Кожа (потовые железы) Амнион
Почки Фаллопиевы трубы
Мочевой пузырь Слизистая оболочка матки
Паренхиматозные клетки печени Тимус
Класс 3. Отсутствие местного синтеза IgA и секреторного компонента
Мочеточник Влагалище
Пищевод Слизистая оболочка рта
В слизистых оболочках индуктивная и продуктивная фазы гуморального иммунного ответа разобщены пространственно в большей степени, чем в других отделах иммунной системы.
Клеточные и гуморальные превращения, связанные с появлением IgA во внешних секретах, представлены на рисунке 18.
В молоко и, по-видимому, в другие внешние секреты молекулы IgA поступают из двух главных источников. Большая часть IgA, выделяемого со слюной, секретами слезной и молочной желез, а также пищеварительного и респираторного трактов, образуется плазматическими клетками. Однако IgA, обнаруживаемые в различных внешних секретах, могут иметь и системное происхождение. Продуцируясь клетками слизистых оболочек одних органов, они поступают в кровь и переносятся в слизистые покровы других органов.
Антиген после проникновения внутрь пейеровой бляшки активирует Т- и В- лимфоциты, которые по лимфатическому протоку поступают в мезентериальный лимфатический узел, а затем в кровь, селезенку, снова в кровь и селективно локализуются в лимфатических образованиях всех слизистых оболочек и экзо-
Плазматическая
клетка
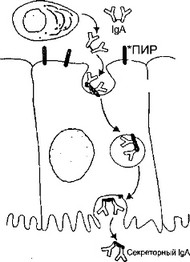
Рис. 18. Синтез и транспорт IgA через эпителий слизистой оболочки.
*ПИР — полииммуноглобулиновый рецепт [1523]
кринных секреторных железах — молочных, слюнных и слезных (рис. 19). При этом Т-лимфоциты локализуются преимущественно между эпителиальными клетками слизистой оболочки, обеспечивая клеточный иммунный ответ, а В- лимфоциты — в lamina propria, где они дифференцируются в плазматические клетки и синтезируют IgA. Около 90% плазматических клеток в lamina propria продуцируют IgA, в то время как в лимфатических узлах доля таких клеток составляет всего 2—5% [139].
Уникальна в этом отношении роль печени. Имеются убедительные данные, свидетельствующие о том, что гепатоциты избирательно связывают и в последующем транспортируют IgA в желчь, тем самым усиливая систему секреторного IgA кишечника [420].
Возможной функцией печени также является удаление комплексов антиген - IgA из циркулирующей крови с желчью в кишечник. Изучение противовирусного иммунитета убедительно продемонстрировало, что степень защиты респираторного и желудочно-кишечного трактов от локальных вирусных инфекций прямо коррелирует с уровнем специфических секреторных IgA, а не с уровнем
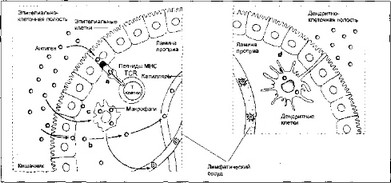
Рис. 19. Пути преодоления эпителиального барьера кишечника «алиментарными» антигенами [Mayer L., Shaul L. Nature Rev. Immunol. 2004, 4, 409]: a — с помощью МНС-комплекса эпителия кишечника;
Ь — абсорбцией капиллярами; с — поглощением макрофагами;
d — поглощением дендритными клетками с последующим поступлением в лимфатические узлы.
антител сыворотки крови. В основе противовирусного действия секреторного IgA лежит инактивация вируса [420].
Стабильная структура, выраженный аффинитет к поверхности слизистых оболочек, преобладающее содержание в секрете молочной железы обусловливают биологическую роль секреторного IgA в защите организма от неблагоприятного воздействия различных патогенных агентов, в том числе вирусов.
IgA синтезируется в димерной форме в клетках lamina propria и после связывания с иммуноглобулиновым рецептором, синтезированным в эпителиальных клетках, транспортируется на поверхность слизистой оболочки. В момент выхода IgA в просвет кишечника рецептор частично расщепляется, в результате чего в составе IgA остается фрагмент рецептора, который называют секреторным компонентом (рис. 18). Таким образом, секреторный IgA является продуктом кооперации двух типов клеток — плазматических и эпителиальных.
Секреторный IgA образуется не только в димерной, но и в тетрамерной форме, что усиливает его вируснейтрализующую способность. Секреторный компонент предохраняет IgA от расщепления протеолитическими ферментами, что обусловливает его значительные преимущества перед антителами других классов. Секреторный IgA нейтрализует вирус не только в просвете кишечника, но и при транспортировке его внутрь клетки. Димер IgA может нейтрализовать вирус в подслизистой оболочке кишечника, а затем, связавшись с рецептором, транспортировать его в просвет кишечника [139].
Димерные иммуноглобулины класса А (11S IgA), соединенные посредством j-цепи в единую структуру с секреторным компонентом, представляют собой
уникальный пример эволюционной адаптации иммуноглобулинов на слизистых покровах для эффективного функционирования в условиях постоянного воздействия антигенов различной природы.
Исходя из способности к локальному синтезу молекул секреторного компонента, а также димерной формы IgA-местно расположенными плазматическими клетками, ткани слизистых покровов можно разделить на три основных класса:
Класс 1. Выраженный местный синтез IgA и секреторного компонента
Желудочно-кишечный тракт Верхний отдел респираторного тракта Носовая полость Среднее ухо Желчный пузырь
Класс 2. Местный синтез секреторного компонента при отсутствии местного синтеза IgA
Кожа (потовые железы) Амнион
Почки Фаллопиевы трубы
Мочевой пузырь Слизистая оболочка матки
Паренхиматозные клетки печени Тимус
Класс 3. Отсутствие местного синтеза IgA и секреторного компонента
Мочеточник Влагалище
Пищевод Слизистая оболочка рта
В слизистых оболочках индуктивная и продуктивная фазы гуморального иммунного ответа разобщены пространственно в большей степени, чем в других отделах иммунной системы.
Клеточные и гуморальные превращения, связанные с появлением IgA во внешних секретах, представлены на рисунке 18.
В молоко и, по-видимому, в другие внешние секреты молекулы IgA поступают из двух главных источников. Большая часть IgA, выделяемого со слюной, секретами слезной и молочной желез, а также пищеварительного и респираторного трактов, образуется плазматическими клетками. Однако IgA, обнаруживаемые в различных внешних секретах, могут иметь и системное происхождение. Продуцируясь клетками слизистых оболочек одних органов, они поступают в кровь и переносятся в слизистые покровы других органов.
Антиген после проникновения внутрь пейеровой бляшки активирует Т- и В- лимфоциты, которые по лимфатическому протоку поступают в мезентериальный лимфатический узел, а затем в кровь, селезенку, снова в кровь и селективно локализуются в лимфатических образованиях всех слизистых оболочек и экзо-
Плазматическая
клетка
Рис. 18. Синтез и транспорт IgA через эпителий слизистой оболочки.
*ПИР — полииммуноглобулиновый рецепт [1523]
кринных секреторных железах — молочных, слюнных и слезных (рис. 19). При этом Т-лимфоциты локализуются преимущественно между эпителиальными клетками слизистой оболочки, обеспечивая клеточный иммунный ответ, а В- лимфоциты — в lamina propria, где они дифференцируются в плазматические клетки и синтезируют IgA. Около 90% плазматических клеток в lamina propria продуцируют IgA, в то время как в лимфатических узлах доля таких клеток составляет всего 2—5% [139].
Уникальна в этом отношении роль печени. Имеются убедительные данные, свидетельствующие о том, что гепатоциты избирательно связывают и в последующем транспортируют IgA в желчь, тем самым усиливая систему секреторного IgA кишечника [420].
Возможной функцией печени также является удаление комплексов антиген - IgA из циркулирующей крови с желчью в кишечник. Изучение противовирусного иммунитета убедительно продемонстрировало, что степень защиты респираторного и желудочно-кишечного трактов от локальных вирусных инфекций прямо коррелирует с уровнем специфических секреторных IgA, а не с уровнем
Рис. 19. Пути преодоления эпителиального барьера кишечника «алиментарными» антигенами [Mayer L., Shaul L. Nature Rev. Immunol. 2004, 4, 409]: a — с помощью МНС-комплекса эпителия кишечника;
Ь — абсорбцией капиллярами; с — поглощением макрофагами;
d — поглощением дендритными клетками с последующим поступлением в лимфатические узлы.
антител сыворотки крови. В основе противовирусного действия секреторного IgA лежит инактивация вируса [420].