
В течение многих лет разрабатывались методы выращивания небольших количеств клеток животных в лабораторных условиях. Однако наладить массовое культивирование таких клеток оказалось не простой задачей. Способ выращивания, характер используемой среды, методы управления и контроля в значительной степени зависят от типа выращиваемых клеток. Все культивируемые клетки первоначально получают от животных путем механической или ферментативной дезагрегации нормальных или малигинизированных тканей либо с помощью перфузии in vivo. Тканевые эксплантаты или клетки, помещенные в подходящую среду, развиваются в первичные культуры с «нормальным» (диплоидным) или «ненормальным» (трансформированным или происходящим из опухоли) карио- типом. Классификация культивируемых клеток представляет определенные трудности. Различают первичные культуры клеток и клеточные линии [1375]. В производстве вакцин используют, главным образом, три типа клеток; первичные; диплоидные клеточные линии; непрерывные (постоянные) клеточные линии. Первичная культура — это культура, происходящая от клеток тканей или органов, взятых непосредственно из организма. Культура считается первичной до тех пор, пока ее не субкультивируют, после чего она становится клеточной линией.
Первичные культуры обычно получают трипсинизацией тканей куриных эмбрионов или тканей (чаще всего почек), взятых от других видов здоровых животных. Возраст используемых эмбрионов может значительно различаться и влиять на выход и жизнеспособность клеток. Для более полного контроля доноров ткани их следует брать из SPF-хозяйств.
Первичные культуры клеток почек зеленых мартышек обычно используют для приготовления полиовирусной вакцины. Поскольку обезьяны являются очень дорогим объектом для увеличения выхода клеток почки дезагрегируют методом перфузионной трипсинизации in vivo, а затем первичную культуру выращивают на микроносителях. Для изготовления других медицинских вакцин широко применяют первичную культуру клеток почки кроликов, телят и других животных. Для изготовления вакцин, используемых в ветеринарной медицине, чаще всего применяют первичные культуры клеток куриных эмбрионов и почек естественного хозяина или восприимчивых животных. Первичные культуры в зависимости от типа ткани и условий выращивания значительно различаются. Некоторые из них погибают через несколько дней после посева, тогда как другие могут длительно сохраняться в культуре без заметных морфологических и биохимических изменений. С момента приготовления первичной культуры до остановки роста клеток и гибели культуры после серийных пересевов проходит от нескольких недель до нескольких месяцев. Клетки эмбрионального происхождения при культивировании обычно сохраняют жизнеспособность более продолжительное время.
Линия клеток — первичная культура, со времени получения субкультуры клеточные линии могут иметь ограниченный срок жизни in vitro (например, диплоидные фибробласты человека и животных), а могут размножаться in vitro неогра
ниченно долго. В последнем случае их называют постоянными, стабильными или непрерывными клеточными линиями.
Линия диплоидных клеток - клеточная линия, в которой, по крайней мере, 75% клеток обладает кариотипом нормальных клеток исходного вида. Следует отметить, что диплоидный набор хромосом не обязательно эквивалентен диплоидному кариотипу, поскольку существуют ситуации, при которых клетка может терять один тип хромосом и приобретать другой. Следовательно, кариотип клетки изменился, а диплоидный набор хромосом сохранился. Такие клетки следует считать «псевдодиплоидными». Диплоидные культуры фибробластов человека могут пассироваться многократно. Такие культуры часто называют штаммами. Некоторые из них могут сохранять диплоидный статус в течение 80 и даже более популяционных делений.
В связи с трудностями получения соответствующих количеств первичных культур клеток почки зеленой мартышки (по причине стоимости) и преимуществами использования предварительно изученных клеточных культур для производства вакцины против полиомиелита и других вакцин были лицензированы диплоидные линии клеток WI-38 и MRC-5. Существуют сомнения в том, что ростовые свойства и продуктивность диплоидных клеток варьируют. Даже хорошо охарактеризованные диплоидные линии клеток ведут себя неодинаково при работе с ними различных исследователей.
Штамм клеток — популяция однородных клеток (по одному или нескольким маркерам), сохраняющая специфические свойства в течение ограниченного периода культивирования. Штамм клеток может происходить из первичной культуры либо от линии клеток, получающих специфические свойства путем селекции или клонирования.
Линия гетероплоидных клеток. Этот термин означает, что клеточная линия имеет менее 75% клеток с диплоидным набором хромосом. Этот термин не означает, что клетки являются злокачественными или способны расти до бесконечности.
Постоянная (непрерывная) клеточная линия — это клетки, способные субкуль- тивироваться вне организма в течение неограниченного количества пассажей. На уровне знаний о поведении культивируемых клеток невозможно определить точку, когда культура становится стабильной, т. е. приобретает способность расти до бесконечности. Только на основе опыта с фибробластоподобными клетками человека показано, что культура должна субкультивироваться, по крайней мере, 70 раз с интервалом примерно в три дня, прежде чем ее можно будет считать постоянной (непрерывной) линией клеток.
Все культуры с неограниченной способностью к размножению («бессмертные культуры») следует называть постоянными (непрерывными) или стабильными линиями. В вирусологической литературе широко распространен термин «перевиваемые линии клеток», под которым имеются в виду линии клеток, способные к бесконечному росту вне организма [2].
Под трансформацией клеток подразумевают необратимые генетические изменения, характеризующиеся изменением ростовых свойств культивируемых
клеток. Трансформация делает клетки бессмертными и менее зависимыми от питательных факторов и геометрических параметров роста. Изменение ростовых свойств связано с изменением транспортировки питательных веществ через клеточную мембрану и является одной из адаптивных особенностей, позволяющих клеткам размножаться в условиях, неблагоприятных для не трансформированных родительских клеток. Изменение клеточной мембраны сопровождается потерей поверхностного белка (150 кД) и изменением антигенных свойств. Основное достоинство трансформированных клеток - способность к бесконечному размножению и улучшенные ростовые свойства. Трансформированные клетки могут образовывать опухоли у иммунологически толерантных животных. По этой причине трансформацию иногда неправомерно приравнивают к злокачественным изменениям [101]. Первичные культуры клеток, линии диплоидных и трансформированных клеток получили широкое применение в вирусологической практике и явились основой создания крупномасштабных систем культивирования клеток и производства вакцин. Это, в свою очередь, потребовало усовершенствования методов культивирования, организации коммерческого производства сред и аппаратуры для культур клеток.
Получение постоянных линий намного расширило исследования в области клеточной биологии и вирусологии. На их основе появилась возможность получения мутантных и гибридных линий клеток, что открыло перспективу создания новых субстратов, пригодных для культивирования вирусов. Нормальные клетки удается субкультивировать вне организма в течение ограниченного количества пассажей, после чего их размножение прекращается и они, как правило, погибают [78]. Это обычно происходит примерно после 50 генераций, если клетки получены из тканей взрослого организма. Лишь иногда они способны продолжать размножаться в культуре неограниченно долго, превращаясь в так называемые трансформированные постоянные клеточные линии [525]. Трансформация клеток в культуре имеет полиэтиологическую природу. Спонтанная трансформация культур клеток нормальных тканей — относительно редкое событие. Частоту таких трансформаций можно увеличить, обработав клетки мутагенами или некоторыми вирусами.
Все постоянные клеточные линии, в отличие от культур нормальных клеток с ограниченным сроком жизни, являются трансформированными, т. е. измененными по ряду признаков, главным из которых является «бессмертие». Способность к неопределенно длительному размножению в культуре, как правило, связана с анеуплоидным кариотипом. Некоторые линии клеток (например, фиб- робластов хомячка — ВНК-21) хотя и обладают правильным диплоидным числом хромосом, но их кариотип, по-видимому, изменен. Другие критерии трансформации (онкогенность, контактное торможение требования к среде) проявляются у различных линий неодинаково [78]. Все они получены в результате спонтанной или индуцированной трансформации клеток в организме или в культуре. Многие происходят из опухолей животных, хотя не всегда при эксплантации опухолевых клеток in vitro образуются клеточные линии. Линии кле
ток, полученные непосредственно из опухолевых тканей (т.е. трансформированных в организме), часто сохраняют специализированные функции, например, способность продуцировать некоторые ферменты в течение более продолжительного периода, чем клетки, трансформированные в культуре различными методами (физическими, химическими, вирусными агентами). Частота получения постоянных линий у различных видов млекопитающих неодинакова. Клетки мышей подвергались спонтанной трансформации значительно чаще, чем клетки человека или других млекопитающих. Указанные различия, по-видимому, связаны с неодинаковой стабильностью кариотипа разных видов животных [168].
Субкультивирование клеток, а затем и становление постоянной клеточной линии, были более успешными, когда первичную однослойную культуру получали путем посева механически измельченной ткани, чем из суспензии трипси- низированных клеток. Возможно, это связано с уменьшением количества белка в поверхностных структурах клеток в результате обработки трипсином. Для трансформации клеток часто используют вирусы. Клетки почки новорожден-
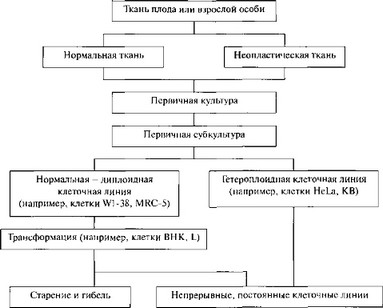
Рис. 13. Схематическое изображение «ответвления» (дивергенции) при культивировании in vitro соматических клеток [52].
ного хомяка (ВНК), например, легко трансформируются вирусом полиомы [1458] или вирусом SV-40, а клетки эмбриона хомяка — герпесвирусом типа 3 лошадей [1468]. В последнем случае клеточные линии экспрессировали вирус - специфические белки, но не продуцировали вирусного потомства. Трансфекция культуры клеток почки эмбриона человека онкогеном вируса SV-40 увеличивала период размножения in vitro от 15-20 до 70-90 поколений. Из популяции таких клеток удалось выделить две постоянные линии, практически не имеющие признаков трансформации [209]. В результате трансфекции гепато- цитов нормальных крыс ДНК вируса SV-40 получена серия клеточных линий SV-40 гепатоцитов [952]. С помощью рекомбинантного вектора SV-40-аденови- руса получены различающиеся по свойствам постоянные линии клеток стромы костного мозга человека, экспрессирующие большой Т-антиген вируса SV-40 [223]. Т-штамм вируса ретикулоэндотелиоза кур вызывал трансформацию культур клеток печени, селезенки, костного мозга, а также тимуса куриных эмбрионов и молодых цыплят. Подавляющее большинство трансформированных линий клеток секретировало вирус, вызывающий летальный ретикулоэндотелиоз у 1—2-дневных цыплят [1371].
Предпочтительными сайтами интеграции вирусного генома оказались ломкие участки в геноме млекопитающих, что обусловлено, по-видимому, их структурными и функциональными особенностями. Результатом взаимодействия вирусного и клеточного геномов могут явиться хромосомные аномалии и приобретение клетками способности к неограниченному размножению.
При длительном культивировании клеточные линии могут изменять те или иные свойства, поэтому для поддержания исходных свойств расплодки клеток, их хранят при низкой температуре и периодически берут для размножения и практического использования. Например, в процессе многих циклов выращивания клеток ВНК-21 в однослойной культуре могут значительно изменяться их первоначальные свойства. Нормальный равномерный рост фибробластоподоб- ных клеток может смениться ростом эпителиоподобных клеток, образующих слои и грозди. От свойств сыворотки также зависит скорость роста клеток и чувствительность к разным типам и штаммам вируса ящура [1610].
Большинство таких изменений характерно для трансформированных клеток других линий.
В отличие от первичных культур клеток и штаммов диплоидных клеток, спектр вирусной чувствительности вновь полученных постоянных линий часто непредсказуем. В настоящее время в мировой практике используется большое количество постоянных линий и создаются новые.
В американской коллекции типовых культур хранятся сотни клеточных линий более чем 50 видов животных. Только из ткани клещей получено более 20 постоянных линий клеток, представляющих интерес для культивирования арбо- вирусов.
Некоторые минимально трансформированные линии иногда называют псевдонормальными, поскольку по всем основным ростовым характеристикам
(выраженная зависимость пролиферации от ростовых факторов и от плотности популяции) эти клетки не отличаются от нормальных. Такие культуры, по-видимому, могут представлять особый интерес в качестве субстратов для выращивания вирусов при изготовлении живых вакцин.
Специфической разновидностью стабильных клеточных линий являются культуры лимфобластоидных и миеломных клеток. Они, как правило, являются суспензионными. Клетки имеют округлую форму, размножаются, не прикрепляясь к стенкам культуральных сосудов, в стационарной культуре не образуют агрегатов [2, 78]. Длительной селекцией по признаку адгезивности из линии клеток плазмоцитомы (линия МОРС 173) получены две линии, представленные фи- бробластоподобными и эпителиовидными клетками [78]. С момента первого сообщения о получении стабильной линии лимфобластоидных клеток количество таких линий быстро возрастало и в настоящее время исчисляется многими сотнями. Некоторые из них нашли применение в клеточной биотехнологии.
Вирус Эпштейна-Барр легко вызывает трансформацию В-лимфоцитов человека, превращая их в стабильные линии клеток. Трансформирующая активность отдельных штаммов вируса исключительно высока и достигает 90—100% [121].
Лимфобластоидные клетки отличаются некоторыми особенностями они просты в обращении, сохраняют диплоидный или псевдодиплоидный кариотип, размножаются с высокой скоростью, в том числе, в крупномасштабных культурах. Их линии могут обладать выраженной фагоцитарной активностью [605]. До недавнего времени почти все лимфобластоидные линии клеток человека характеризовались как клетки В-типа и содержали генетическую информацию вируса Эпштейна-Барр, даже если не продуцировали его антигенов. В дальнейшем были получены лимфобластоидные клетки T-типа. Ряд лимфобластоидных линий клеток был получен из лимфом цыплят и индюшат. Их, в отличие от лимфобластоидных клеток человека, получали с большими трудностями, они имели характеристику клеток T-типа. Они обнаруживали нормальный кариотип, присутствие генома вируса болезни Марека, большинство из них продуцировало этот вирус (в небольшом количестве) и при введении цыплятам вызывало развитие типичных лимфом [163]. Вирус болезни Марека хорошо размножался в линии лимфобластоидных клеток MS В1-41 С, полученной от инфицированных кур [783]. Из культуры клеток костного мозга индеек путем трансформации вирусом миелоцитоматоза (штамм МС-31) получена постоянная линия клеток LSTC- SF2, которая после 450 пассажей продуцировала вирус МС-31 [839]. Лимфобла- стоидная клеточная линия была получена из клеток лимфомы селезенки кроликов, зараженных вирусом герпеса Саймири [211], а также из клеток тимуса крупного рогатого скота при спорадическом лейкозе [308]. Несколько линий лимфобластоидных клеток получены из тимуса и лимфоцитов цереброспинальной жидкости крупного рогатого скота и оленя, зараженных вирусом злокачественной катаральной лихорадки овечьего типа. Линия лимфобластоидных клеток оленя продуцировала инфекционный вирус и вызывала заболевание кроликов. Линии клеток, полученные от крупного рогатого скота, этим свойством не обла
дали. Линия лимфобластоидных клеток SBLC-1, полученная из опухоли кожного лейкоза коровы, имела Т-клеточное происхождение, не содержала интегрированного провируса лейкоза и не экспрессировала вирусный антиген. Т-лимфоб- ластоидная линия клеток кошки (МУА-1) оказалась высокочувствительной к вирусу иммунодефицита кошек [1000]. Культуры лимфобластоидных клеток человека обнаруживали чувствительность ко многим вирусам. Их часто использовали в качестве субстрата при культивировании вирусов иммунодефицита человека (ВИЧ-1, ВИЧ-2). Т-лимфобластоидные клетки человека (линия Н9) сохранили чувствительность к вирусу (ВИЧ-1, ВИЧ-2) и ростовые свойства родительских клеток после адаптации к бессывороточной среде Chilton, вместо среды RPMI 1640 с 10% сыворотки эмбрионов крупного рогатого скота [1691]. Гемато- поэтические клеточные линии человека использовались для размножения гер- песвируса-6 человека и других вирусов. Наконец, клеточные линии, происходящие из миелом и сохранившие способность синтезировать иммуноглобулины, могут служить основой быстро развивающегося биотехнологического способа получения моноклинальных антител.